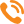Медоносная пчела занимает широкий ареал – от экватора до Северного полярного круга, что связано с использованием ею большого комплекса этолого-физиологических адаптаций, которые можно дифференцировать на индивидуальные и социальные. Их роль меняется в зависимости от ситуации, чем достигается высокая мобильность биологически целесообразного реагирования, обеспечивающая медоносной пчеле широкую экологическую валентность.
Медоносная пчела занимает широкий ареал – от экватора до Северного полярного круга, что связано с использованием ею большого комплекса этолого-физиологических адаптаций, которые можно дифференцировать на индивидуальные и социальные. Их роль меняется в зависимости от ситуации, чем достигается высокая мобильность биологически целесообразного реагирования, обеспечивающая медоносной пчеле широкую экологическую валентность.
Преодоление медоносной пчелой длительной зимовки в условиях умеренного и холодного климата основано на использовании большого комплекса индивидуальных и социальных этолого-физиологических адаптаций. Их приобретение и/или совершенствование сопряжено с эволюцией социальности (Е.К.Еськов, 1992, 1995, 2003). На этом генеральном направлении в филогенезе вида доминирующее значение приобрели приспособления, обеспечивающие накопление кормовых запасов, их экономное использование в течение зимовки и регуляцию внутригнездовой температуры.
Лабильный механизм внутригнездовой терморегуляции работает на базе генетически детерминированного изменения соответственно биологической ситуации активности термогенеза и интенсивности тепловых потерь. Эффективность функционирования этого механизма связана с изменчивостью плотности агрегирования пчел, стимулируемого охлаждением, вокруг исходно нагретой зоны гнездового пространства, которое становится тепловым центром (Е.К.Еськов, 1995). Плотность агрегирования и соответствующее этому уменьшение объема гнезда, занимаемого пчелами, находятся в прямой зависимости от интенсивности их охлаждения. Поэтому порождаемое охлаждением уменьшение поверхности агрегировавшихся пчел снижает тепловые потери в результате ограничения воздухообмена между тепловым центром и внешней средой. Но из-за этого в нем повышается концентрация СО2 (V.Lacher, 1964), выделяемого пчелами, что возбуждает их. К усилению возбуждения имеет отношение совмещение тепловыми рецепторами функции рецепторов СО2. По указанным причинам при понижении внешней температуры имеет высокую вероятность усиление разогрева теплового центра.
Упорядоченность и лабильность терморегуляторных механизмов, используемых пчелиной семьей позволяют переживать зимовку при минимальных затратах энергии и соответственно кормовых запасов. Значительные колебания внешней температуры стимулируют необходимость коррекции структуры агрегации пчел в гнезде и локализации в межсотовых пространствах. При этом в некоторых ситуациях, порождаемых внутригнездовыми причинами (размещение кормовых запасов, численность пчел в гнезде, их локализация на сотах и др.), возможно инадаптивное реагирование, осложняющее в дальнейшем реализацию пчелами врожденных механизмов терморегуляции, что снижает вероятность успешного преодоления зимовки. Адекватное реагирование пчел на случайные изменения условий зимовки выражается в приобретении ими способности образовывать пространственно-временные структуры, обеспечивающие оптимизацию внутригнездового микроклимата в экстремальных ситуациях. Их вариабельность, очевидно, выступает в роли некоторого упреждающего сигнала, стимулирующего выбор оптимального реагирования на возможные изменения широкого спектра возможных колебаний внешней температуры. В этом выражается способность пчелиной семьи к самоорганизации, спонтанному образованию и развитию упорядоченности функционирования ее структурных единиц — отдельных особей и их скоплений.
Агрегирование не одномоментный процесс, связанный с понижением температуры. Начиная с 20...16°С пчелы спорадически образуют небольшие временные скопления на сотах с кормом. По мере понижения температуры до уровня, стимулирующего холодовое оцепенение, активность агрегирования пчел возрастает. По наблюдениям О.С.Львова (1964), пчелы небольших семей агрегируются при 13°С, средних – при 7°С, в больших — при 5°С. В опытах I.B.Free, L.Spenser-Booth (1959), выполненных на изолированных группах пчел, агрегирование 28% пчел происходило при 20°С, 77% – при 15°С и 100% — при 10°С.
 На локализацию в гнезде теплового центра агрегирования пчел влияют местоположение летка, распределение меда на сотах и утепление. При этом прослеживаются две несвязанные тенденции: пчелы стремятся занять пустые ячейки и верхнюю более теплую часть гнезда. Поэтому нередко с осени пчелы локализованы в верхней части гнезда, смещенной к передней стенке улья, занимая не только пустые, но и медовые соты (рис. 1).
На локализацию в гнезде теплового центра агрегирования пчел влияют местоположение летка, распределение меда на сотах и утепление. При этом прослеживаются две несвязанные тенденции: пчелы стремятся занять пустые ячейки и верхнюю более теплую часть гнезда. Поэтому нередко с осени пчелы локализованы в верхней части гнезда, смещенной к передней стенке улья, занимая не только пустые, но и медовые соты (рис. 1).
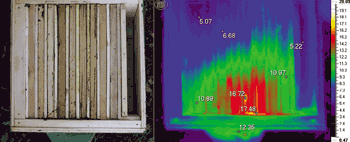
В смежных межсотовых пространствах локализуется неодинаковое количество пчел, а их плотность зависит от температуры в разных зонах гнезда. Поскольку наибольшему охлаждению подвергается нижняя часть межсотового пространства, то плотность агрегации пчел в этой зоне достигает наибольшего значения (рис. 2). Плотность пчел минимальна в предпочитаемой ими температурной зоне, здесь они могут свободно передвигаться. Над этой зоной в соответствии с температурным градиентом плотность агрегации выше, чем в центре, но ниже, чем в нижней части локализации пчел. Таким образом, неоднородная агрегация пчел обусловлена внешней температурой и зависит от наличия в гнезде разогретой зоны.
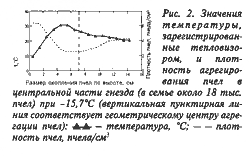 Изменения температуры в самой теплой зоне гнезда и над ее поверхностью (непосредственно под холстиком) в зависимости от внешней температуры представлены на рис. 3. Эта зависимость имеет выраженную тенденцию к ослаблению от начала к завершению зимовки. Причем в конце зимовки связь между колебаниями внешней и внутригнездовой температурами совсем не прослеживается. Так, в ноябре коэффициент корреляции между указанными температурами находился на высоком отрицательном уровне (r = –0,81±0,10), а в марте изменился на слабоположительный (r = 0,13±0,04).
Изменения температуры в самой теплой зоне гнезда и над ее поверхностью (непосредственно под холстиком) в зависимости от внешней температуры представлены на рис. 3. Эта зависимость имеет выраженную тенденцию к ослаблению от начала к завершению зимовки. Причем в конце зимовки связь между колебаниями внешней и внутригнездовой температурами совсем не прослеживается. Так, в ноябре коэффициент корреляции между указанными температурами находился на высоком отрицательном уровне (r = –0,81±0,10), а в марте изменился на слабоположительный (r = 0,13±0,04).
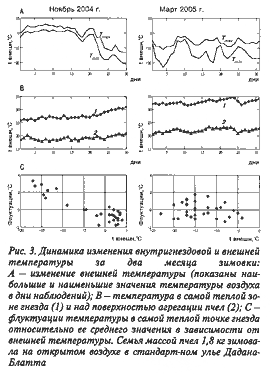 В начале зимовки (октябре—ноябре) диапазон флуктуаций внутригнездовой температуры выше таковой на ее завершающих фазах. Это связано с отличиями в интенсивности реагирования пчел на колебания внешней температуры. Одни и те же пчелы в начале зимовки активнее, чем перед ее окончанием, реагируют на понижения внешней температуры интенсификацией термогенеза. Диапазон флуктуаций внутригнездовой температуры начинает уменьшаться с приближением к завершению первой половины периода зимовки (в середине декабря) и достигает минимального значения в марте (рис. 3.). Исходя из этого, степень дисперсии (вариабельности) температурных флуктуаций может служить одним из показателей физиологического резерва пчелиной семьи, то есть чем больше дисперсия, тем выше число возможных состояний и различных типов перехода к ним, — эффективнее работает механизм температурной адаптации.
В начале зимовки (октябре—ноябре) диапазон флуктуаций внутригнездовой температуры выше таковой на ее завершающих фазах. Это связано с отличиями в интенсивности реагирования пчел на колебания внешней температуры. Одни и те же пчелы в начале зимовки активнее, чем перед ее окончанием, реагируют на понижения внешней температуры интенсификацией термогенеза. Диапазон флуктуаций внутригнездовой температуры начинает уменьшаться с приближением к завершению первой половины периода зимовки (в середине декабря) и достигает минимального значения в марте (рис. 3.). Исходя из этого, степень дисперсии (вариабельности) температурных флуктуаций может служить одним из показателей физиологического резерва пчелиной семьи, то есть чем больше дисперсия, тем выше число возможных состояний и различных типов перехода к ним, — эффективнее работает механизм температурной адаптации.
Повышение в ходе зимовки независимости внутригнездовой температуры от внешней указывает на развитие у пчел специфических приспособлений к ее вариабельности. Приспосабливаясь к изменениям внешней температуры, они стремятся к автономизации. В результате открытая система — пчелиная семья приобретает некоторую независимость от флуктуаций фактора, влияющего на состояние зимующих особей. Подтверждением этому служит сильная корреляционная связь между температурами в самой теплой зоне гнезда и верхней частью локализации пчел (r = 0,82±0,11).
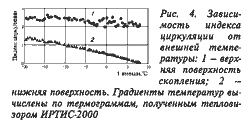 Индекс циркуляции тепла в скоплении пчел (отношение наружного градиента температуры к внутреннему) неодинаков в разных направлениях, что зависит от интенсивности тепловых потоков из внутренних зон к внешним. С уменьшением внешней температуры индекс циркуляции тепла на нижней поверхности увеличивается, а на верхней — меняется незначительно (рис. 4).
Индекс циркуляции тепла в скоплении пчел (отношение наружного градиента температуры к внутреннему) неодинаков в разных направлениях, что зависит от интенсивности тепловых потоков из внутренних зон к внешним. С уменьшением внешней температуры индекс циркуляции тепла на нижней поверхности увеличивается, а на верхней — меняется незначительно (рис. 4).
Если теплоизолирующие свойства воздуха, окружающего агрегировавшихся пчел, постоянны, то изменение индекса циркуляции тепла будет характеризовать степень физиологических изменений теплоизоляционных свойств пчел, примыкающих к периферии скопления. Чем меньше разность Тц – Тпов (Тц и Тпов соответственно температуры в самой теплой точке и на поверхности скопления пчел), тем меньше энергии тратят они на поддержание теплового гомеостаза внутри своего скопления. За весь период зимовки разность в направлении к верхней поверхности меняется незначительно и не превышает 12°С. Ее минимизации способствует герметизация пчелами всех щелей в верхней части гнезда. В то время как тепловое поле на нижней поверхности меняется постоянно, а разность Тц – Тпов может достигать 28°С из-за высокого значения температуры оцепенения пчел и необходимости поддержания в связи с этим положительной температуры на периферии их агрегирования (поверхности скопления). Внешний градиент температур также максимален на стороне нижней поверхности скопления пчел. Поэтому основная регуляторная функция, способствующая поддержанию температурного гомеостаза, обеспечивается пчелами, локализующимися ниже теплового центра. При понижении внешней температуры тепловой центр смещается к зоне соприкосновения пчел с холодным воздухом, на что указывает увеличение температурного градиента (см. рис. 2). В результате интенсивность теплового потока к пчелам, образующим нижнюю часть периферической области их скопления, возрастает.
Распределение тепловых полей указывает на отсутствие четко выраженной стабильности в распределении тепловой картины внутри гнезда, как это принято изображать в виде изотерм. В действительности чередования периодов относительного покоя и активной теплогенерации пчел существенно дестабилизирует температурный режим в различных зонах их локализации. Этому способствуют миграции пчел внутри образуемого ими скопления. С понижением внешней температуры доля активных пчел, непосредственно примыкающих к нижней поверхности их скопления, возрастает (Е.К.Еськов, В.А.Тобоев, 2006).
Таким образом, в скоплении зимующих пчел нет единого центрального механизма, способного контролировать и регулировать внутригнездовую температуру. По-видимому, отдельно взятые особи ведут себя как простые объекты с малым числом эффективных степеней свободы. Но каждая пчела внутри скопления в зависимости от местоположения и функциональных возможностей может находиться в неодинаковых условиях, чему соответствует их разное участие в теплорегуляции занимаемого гнездового пространства. Особи, локализующиеся в теплой части гнезда, имеют относительно постоянную температуру тела. Они, вероятнее всего, не участвуют в активной термогенерации. Охлаждение нижней части скопления стимулирует активное тепловыделение преимущественно посредством микровибраций крыловой мускулатуры пчел непрямого действия. Некоторая часть насекомых, образующих периферическую часть скопления, может находиться в состоянии неглубокого холодового оцепенения. Случайно дифференцируясь по локализации в скоплении, пчелы образуют упорядоченную динамическую структуру. Ее динамичность, выражающаяся в миграциях пчел, определяется потребностью в пополнении ими индивидуальных кормовых запасов и вариабельностью внешней температуры.
Е.К.ЕСЬКОВ,
В.А.ТОБОЕВ
Российский государственный аграрный заочный университет,
Чувашский государственный университет



















 Адрес редакции журнала "Пчеловодство":
Адрес редакции журнала "Пчеловодство":