Тепловой режим пчелиных семей интересует пчеловодов со времен перехода к стационарному пчеловодству и вплоть до сегодняшнего дня. Родоначальником исследований температурного режима семей по праву считается R.A.F.Reamur (1734). Используя изобретенный им термометр, ученый наблюдал в зимнем клубе пчел температуру 12…19oС.
Тепловой режим пчелиных семей интересует пчеловодов со времен перехода к стационарному пчеловодству и вплоть до сегодняшнего дня. Родоначальником исследований температурного режима семей по праву считается R.A.F.Reamur (1734). Используя изобретенный им термометр, ученый наблюдал в зимнем клубе пчел температуру 12…19oС.
Однако первые же исследователи столкнулись с большим разбросом полученных данных в гнезде как летом, так и зимой. Они отмечали показатель температуры от 10…12 до 35oС и более. И лишь к концу XIX века стало вполне очевидно, что температура зимнего и летнего гнезда неоднородна как во времени, так и по объему.
С внедрением в практику измерений термопар и других малогабаритных датчиков появилась возможность проводить непрерывные наблюдения за показателями температуры во многих точках гнезда. E.F.Phillips и G.S.Demuth (1914), используя 19 датчиков, установили, что как только температура достигнет 13,9oС — образуется клуб. Если наружная температура продолжает снижаться, клуб становится все более и более сжатым, а при потеплении распадается.
В дальнейшем изучать температурный режим семей начали отечественные и зарубежные исследователи (A.Budel, 1949; Т.С.Жданова, 1963; С.D.Owens, 1971). Они установили, что температурное поле семьи неоднородно, построенные ими изотермы показывают наличие сравнительно небольшой области с температурой 30oС и более. По направлению к поверхности она постепенно снижается примерно до 10oС, а за пределами клуба всегда близка к температуре наружного воздуха.
Наряду с топографией температурного поля большой интерес представляет механизм, посредством которого семьи обеспечивают гомеостаз своего гнезда. По мнению E.F.Phillips и G.S.Demuth (1914), клуб представляет собой агрегацию в виде шара, покрытого снаружи плотным слоем пчел, образующих скорлупу. Как полагали исследователи, она предотвращает потерю тепла, выработанного в центральной части, где пчелы находятся в активном состоянии.
Эта гипотеза была принята пчеловодной общественностью, вошла в учебники и имеет всеобщее распространение. В соответствии с ней основным механизмом регулирования температурного режима зимнего клуба считается расширение-сжатие. При уплотнении уменьшается его пористость, что минимизирует конвективные потоки в нем, а сокращение внешней поверхности уменьшает рассеивание тепла в окружающую среду (C.L.Corkins, 1932; P.Lavie, 1954; и др.).
Однако, как показали О.С.Львов (1967), V.Hallund (1956), В.С.Коптев (1979), в семьях средней силы при –5oС пчелы внутри клуба занимают почти все свободное пространство и их сжатие становится практически невозможным. Способность тружениц улья различать разницу температур в 0,25oС продемонстрирована H.Heran (1952), но механизмы на уровне семьи, которые интегрируют поведение отдельных индивидуумов, до сих пор непонятны.
Гипотезы, пытающиеся объяснить механизм терморегуляции, M.L.Winston (1987) подразделяет на две группы: концепция «суперорганизма» и концепция «отдельной пчелы». В соответствии с первой гипотезой пчелы подчиняют свои действия реакции семьи в целом, но нет единого мнения, кто ее вырабатывает. По E.F.Phillips, эта роль отводится матке и расплоду. Однако несостоятельность таких представлений вполне очевидна. Температурный режим устойчиво поддерживается как в безрасплодных, так и в безматочных семьях.
R.F.A.Moritz, E.E.Southwick (1992) описывают опыт, когда в искусственных экспериментальных условиях небольшое количество расплода располагалось в одном секторе сота, где собирались пчелы, а пища находилась на некотором расстоянии. Особей было недостаточно, чтобы одновременно обеспечить обе зоны. При низкой температуре пчелы были вынуждены выбирать между кормом и расплодом. Они неизменно переходили в зону, где располагалась пища, и допускали застывание расплода. Так же пчелы поступали, когда следовало делать выбор между зафиксированной на соте маткой и кормом. В условиях стресса, являющегося следствием низкой температуры, труженицы улья продвигались к меду. Авторы пришли к выводу, что матка не контролирует поведение пчел, связанное с обогревом гнезда, а лишь следует за клубом и продвигается к обогреваемому ядру. Пчелы клуба не идут за маткой, позволяя ей, однако, располагаться в центре. Подвергая семью теплой или холодной камеры, действию феромонов или звуков другой семьи, находящейся в камере с противоположными условиями, изменения терморегуляции также не были замечены.
В соответствии с концепцией «отдельной пчелы» каждая особь действует в собственных интересах, поддерживая температуру своего тела на оптимальном уровне, а терморегуляция на уровне семьи является просто выгодным эффектом суммы поведений индивидуумов. Этот же принцип, по существу, поддерживает и М.Линдауэр (1960), констатируя, что каждая особь ведет себя сообразно с теми локальными условиями гнезда, в которых она оказалась.
Нами предложена версия регулирования микроклимата в гнезде, основанная на теплофизике происходящих процессов. В соответствии с ней семья в целом и зимний клуб в частности представляют собой ограниченный объем, внутри которого действуют источники метаболического тепла, выделяемого особями. Мощность каждого из них невелика и может зависеть от температуры и места, в котором он находится.
В соответствии с законом Фурье температурное поле такого объема формируется в результате баланса всех выделений тепла и его потерь во внешнюю среду и выражается уравнением
Это уравнение должно быть дополнено граничными условиями — температурой теплоотдающей поверхности или интенсивностью ее теплообмена с внешней средой.
Если клуб представить в виде шара и за условие однозначности принять температуру его поверхности, то с рядом упрощающих предпосылок температуру в его центре можно выразить формулой (Г.Карслоу и Д.Егер, 1964)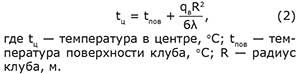
Из этой формулы следует, что температура по направлению к центру всегда растет сама собой и пчелам для создания обычно наблюдаемого в клубе температурного поля нет необходимости прибегать к каким-либо поведенческим действиям. Достаточно собраться вместе, чтобы механизм их обогрева заработал сам собой. В сформированном клубе, как и в обычном гнезде, каждая особь ведет себя в соответствии с локальными условиями, которые ее окружают (М.Линдауэр, 1960). При температуре ниже 20oС они неподвижны, а зону с показателями ниже 13oС покидают. Уходят и из зоны с температурой выше 36oС. Поэтому температура наружной поверхности клуба поддерживается на уровне 13oС. Из формулы (2) также следует, что температурный режим клуба определяется не только его объемом (в формуле через R2), но и теплопроводностью слоя пчел λ и интенсивностью их тепловыделения qв.
Для определения энергетических затрат семьи нами разработан и в течение нескольких лет применяется специальный градиентный улей-калориметр (ж-л «Пчеловодство №4, 2007), позволяющий с высокой точностью регистрировать выделяемую тепловую энергию в условиях, приближенных к нормальной жизнедеятельности. Мы изучили энергетические затраты основных пород пчел, распространенных на территории Российской Федерации, в течение годового цикла. Так, заселенная в улей-калориметр семья среднерусской породы две зимы (2002/03 и 2003/04 гг.) находилась на открытой веранде под навесом. Улей-калориметр оборудовали датчиками, позволяющими определять температуру и влажность внутри и снаружи, для защиты от ветра и механических повреждений его накрыли деревянным кожухом. Сигналы от датчиков по кабелю поступали в лабораторию, где их измеряли и фиксировали.
Датчиком температуры служили стандартные медные термометры в улье с сопротивлением 100 Ом, на улице — 53 Ом. Датчик влажности в улье — резистивный, приводимый в действие от влагочувствительной мембраны. В качестве вторичного использовали комбинированный прибор УППИП-60М, позволяющий измерять как электродвижущую силу (ЭДС), развиваемую батареей термопар калориметра, так и сопротивление термометров. Сигналы от датчика влажности фиксировали с помощью неравновесного моста. Влажность наружного воздуха определяли аспирационным психрометром, с помощью которого был протарирован датчик влажности в улье.
 Ежедневно утром и вечером на протяжении всего периода наблюдений измеряли уровень термогенеза, температуры и влажности в улье над гнездом, а также температуры и влажности наружного воздуха. Измерения термогенеза семьи и температуры наружного воздуха по мере необходимости дополняли непрерывной записью с помощью автоматического потенциометра КСП-2.
Ежедневно утром и вечером на протяжении всего периода наблюдений измеряли уровень термогенеза, температуры и влажности в улье над гнездом, а также температуры и влажности наружного воздуха. Измерения термогенеза семьи и температуры наружного воздуха по мере необходимости дополняли непрерывной записью с помощью автоматического потенциометра КСП-2.
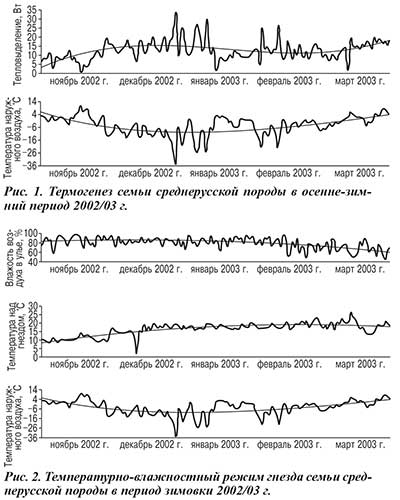
Динамика тепловыделений семьи и температуры внешней среды в осенне-зимний период 2002/03 г. изображена на рисунке 1. Температура внешней среды в этот период изменялась в широком интервале. В начале зимовки она поднималась до 8…11oС, термогенез семьи составлял 3–4 Вт; в конце декабря — опускалась до –25oС, в ночные часы — до –35oС, термогенез возрос до 33,6 Вт.
Если в начале из-за невысокого тепловыделения пчел температура над гнездом составляла 9…12oС, то в связи с вызванным похолоданием ростом термогенеза она возросла до 18…20oС (рис. 2). Относительная влажность воздуха в течение зимовки составляла 60–95%.
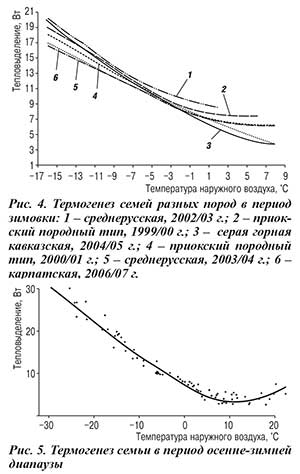
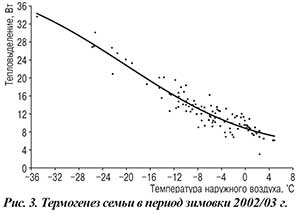 При анализе источников литературы складывается мнение, согласно которому пчелы при понижении наружной температуры сжимают клуб, а при повышении — расширяют, сохраняя постоянство теплообразования семьи. Однако это не соответствует действительности. Полученные нами величины термогенеза семьи сопоставили с температурой наружного воздуха (рис. 3). С понижением внешней температуры, несмотря на образование клуба, термогенез семьи и, как следствие, потребление корма возрастают. Это, впрочем, характерно не только для среднерусских, но и для пчел других пород (рис. 4).
При анализе источников литературы складывается мнение, согласно которому пчелы при понижении наружной температуры сжимают клуб, а при повышении — расширяют, сохраняя постоянство теплообразования семьи. Однако это не соответствует действительности. Полученные нами величины термогенеза семьи сопоставили с температурой наружного воздуха (рис. 3). С понижением внешней температуры, несмотря на образование клуба, термогенез семьи и, как следствие, потребление корма возрастают. Это, впрочем, характерно не только для среднерусских, но и для пчел других пород (рис. 4).
Нам удалось проследить за динамикой термогенеза семьи не только во время зимовки, условно отсчитываемой с 1 ноября, но и в предшествующий ей период, начиная с момента выхода последнего расплода. Результаты показали, что зависимость уровня термогенеза семьи от температуры имеет выраженный минимум, соответствующий температуре 12,3oС (рис. 5). С этой точки кривая монотонно возрастает и других минимумов, в том числе в диапазоне рекомендуемых для зимовников –2…+4oС, не имеет.
Казалось бы, если при 12,3oС термогенез и обмен веществ у пчел минимальны, то эту температуру и следует поддерживать в помещениях для зимовки. Однако практика такую предпосылку отвергает. При 12oС пчелы часто вылетают для освобождения кишечника от каловых масс, в зимовнике они этого сделать не могут, волнуются и поднимают температуру.
 Наши наблюдения позволяют сделать следующие выводы.
Наши наблюдения позволяют сделать следующие выводы.
1. После прекращения медосбора и выхода последнего расплода с понижением внешней температуры термогенез семей среднерусской породы постепенно сокращается, при 12…10oС достигая минимального уровня ~5 (6) Вт. При дальнейшем понижении температуры он неуклонно возрастает до 30 Вт при –30oС.
2. Минимальный уровень термогенеза является одним из критериев благополучной зимовки лишь при размещении на воле в регионах с положительной зимней температурой. При зимовке в помещениях температуру, соответствующую минимальному термогенезу, нельзя принимать за оптимальную.
3. Следует согласиться с авторами, рекомендующими зимовку в помещениях с условиями, соответствующими температуре окончательного образования клуба, а именно: при содержании средних семей — около 0oС, сильных — 0… –2oС. При низкой температуре клуб пчел более устойчив, однако термогенез, а значит и расход кормов возрастают.
4. Существуют три четко выраженных уровня термогенеза семей: с третьей декады апреля по первую декаду августа; с третьей декады апреля и со второй декады августа до начала сентября; с начала сентября до начала зимовки.
5. В осенний период с понижением наружной температуры до 10oС термогенез семьи падает, а при дальнейшем его понижении снова растет, и семья переходит в режим зимовки.
В.И.ЛЕБЕДЕВ,
зам. директора ГНУ «НИИ пчеловодства
Россельхозакадемии»,
доктор сельскохозяйственных наук, профессор
А.И.КАСЬЯНОВ,
зав. сектором информации
и научно-технического обеспечения,
кандидат сельскохозяйственных наук
Аннотация:
Дана интерпретация процессов регулирования микроклимата в пчелиной семье, разработан и испытан улей-калориметр, с помощью которого определены энергетические затраты семьи в течение круглогодового цикла ее жизнедеятельности.
Ключевые слова:
термогенез, зимовка, улей-калориметр.
ЛИТЕРАТУРА:
1. Жданова Т.С. Температурный режим пчелиного гнезда и его изменения в течение года: автореф. дисс… канд. биол. наук. — Казань, 1963.
2. Коптев В.Г. Разведение и содержание пчел в Сибири. — Новосибирск: Западно-Сибирское кн. изд-во, 1979.
3. Львов О.С. Накануне зимовки // Пчеловодство. — 1967. — №10.
4. Moritz R.F.A., Southwick E.E. Bees as Superorganisms, аn Evolutionary Reality. — Berlin: Springer-Verlag, 1992.
5. Owens C.D. The Termology of Wintering Honey Bee Colonies // U.S. Dep. Agr. — №1429. — 1971.
6. Winston M.L. The Biology of the Honey Bee. — London: Harvard University Press, Cambridge, Massachussets, 1987.




















 Адрес редакции журнала "Пчеловодство":
Адрес редакции журнала "Пчеловодство":