Генетическая паспортизация — это получение генетически детерминированных (индивидуальных и/или групповых) характеристик с помощью морфологических и/или молекулярных маркеров. Описание морфологических характеристик селекционного материала — элемент классического генетического анализа и селекционного скрининга, его можно считать первым этапом генетической паспортизации. Второй связан с разработкой и использованием биохимических и молекулярно-генетических маркеров.
В настоящее время проведение генетической паспортизации — актуальная задача современной селекции сельскохозяйственных животных, в том числе и селекции медоносной пчелы.
Стихийная, бесконтрольная межпородная гибридизация пчел, проводившаяся на протяжении последних десятилетий, и отбор по полифакториальным, высоковариабельным в пределах нормы реакции признакам (обычно по признаку медовой продуктивности) привели к разрушению эволюционно сложившихся генетических комплексов отдельных пород и появлению межпородных гибридов с нежелательными фенотипическими признаками и непредсказуемыми комбинациями генетического материала. В результате этих процессов на фоне отсутствия систематического, целенаправленного отбора по генетическим маркерам произошло снижение всех хозяйственно ценных показателей исходных пород, и прежде всего неспецифической устойчивости пчел.
До последнего времени в пчеловодстве для оценки чистопородности использовали, как правило, морфофизиологические критерии: окраску тела, длину хоботка, особенности жилкования крыла, кубитальный индекс, яйценоскость маток и др., то есть показатели в той или иной мере подверженные влиянию абиотических факторов внешней среды. Так, реализация генетически обусловленных показателей яйценоскости маток в значительной мере определяется силой семьи, а недокормленность расплода приводит к уменьшению размера имаго пчелы.
В настоящее время в мировой практике для паспортизации пород и индивидуальной паспортизации сельскохозяйственных животных используют преимущественно ДНК-маркеры, не подверженные влиянию факторов внешней среды. Это ядерные в основном микросателлиты (STR-маркеры) и маркеры митохондриальных ДНК (мтДНК). Использование митохондриальных маркеров имеет ряд преимуществ, связанных с особенностями наследования и организацией митохондриального генома.
Митохондриальный геном — это совокупность генов и некодирующих последовательностей мтДНК. Он представляет собой кольцевую молекулу ДНК размером в среднем 16000–20000 пар нуклеотидов (п. н.). Разнообразие митохондриальных геномов в популяции — отдельный генетический ресурс: митохондриальный генофонд.
♦ Особенностью мтДНК как маркера является ее однородительское наследование: мтДНК передается по материнской линии (от матери к детям) в связи с поведением родительских митохондрий при оплодотворении. Число митохондрий в яйцеклетке на три порядка выше, чем в сперматозоиде, и даже если отдельные отцовские митохондрии попадают в яйцеклетку, то они блокируются на молекулярном уровне, и зигота получает только материнский набор митохондрий (Kaneda et al.,Cummins, 2000). Это обстоятельство приводит к отсутствию рекомбинаций в мтДНК, то есть мтДНК наследуется как единый гаплотип — набор тесно сцепленных локусов (Минченко, Дударева, 1990).
♦ Скорость мутаций в мтДНК примерно в 10–20 раз больше, чем в ядерной ДНК (Brown et al., 1979). В основном это связано с отсутствием эффективных систем репарации и активными окислительно-восстановительными процессами, проходящими в митохондрии.При этом накапливаются преимущественно селективно-нейтральные изменения, вносящие свой вклад в полиморфизм мтДНК (Минченко, Дударева, 1990). Так как рекомбинационная изменчивость в мтДНК отсутствует, то мутационный процесс оказывается единственным путем формирования ее полиморфизма. Сохраняющиеся и накапливающиеся мутации митохондриального генома таким образом оказываются отражением в эволюционной истории вида и отдельной особи.
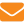
 ♦ Небольшой размер и высокое содержание в клетках мтДНК значительно облегчает исследование ее полиморфизма и позволяет эффективно изучать разнообразие митохондриального генофонда. В настоящее время структуру митохондриального генома активно используют для изучения эволюционных связей и определения породной принадлежности медоносной пчелы и других животных. (Николенко, 2002). В последнем случае изучают первичную структуру маркерной области мтДНК между генами двух субъединиц цитохромоксидазы: субъединицы I и субъединицы II. Известно, что участок, расположенный между ними, образован последовательностью гена тРНКLeu и сложными повторяющимися последовательностями, которые значительно отличаются от остальных участков по нуклеотидному составу Р и Q элементов (рис. 1). Элемент Р этих повторов длиной 54 п. н. состоит только из АТ-пар, а содержание этих нуклеотидов в элементе Q составляет 93% (Cornuet et al., 1991).
♦ Небольшой размер и высокое содержание в клетках мтДНК значительно облегчает исследование ее полиморфизма и позволяет эффективно изучать разнообразие митохондриального генофонда. В настоящее время структуру митохондриального генома активно используют для изучения эволюционных связей и определения породной принадлежности медоносной пчелы и других животных. (Николенко, 2002). В последнем случае изучают первичную структуру маркерной области мтДНК между генами двух субъединиц цитохромоксидазы: субъединицы I и субъединицы II. Известно, что участок, расположенный между ними, образован последовательностью гена тРНКLeu и сложными повторяющимися последовательностями, которые значительно отличаются от остальных участков по нуклеотидному составу Р и Q элементов (рис. 1). Элемент Р этих повторов длиной 54 п. н. состоит только из АТ-пар, а содержание этих нуклеотидов в элементе Q составляет 93% (Cornuet et al., 1991).
Структура повторов оказалась специфична для подвидов. У темной лесной пчелы исследуемый участок представлен набором фрагментов PQQ длиной около 600 пар нуклеотидов. Он включает 3’-область гена цитохромоксидазы I, ген тРНКLeu, Р-элемент, Q-элемент, Q-элемент и 5’-конец гена цитохромоксидазы II (по Никонорову и др., 1988).
В литературе описано всего несколько случаев более длинных вариантов этой области — PQQQ набор фрагментов (по Николенко, 2003). У пчел южных рас, серой горной кавказской, карпатской и других фрагмент Q представлен только одной копией, а длина изучаемой области мтДНК в результате этого составляет около 350 п. н.
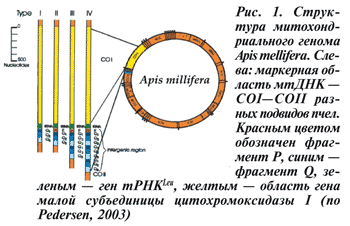
 Авторы этой статьи в целях паспортизации использовали 24 семьи из коллекции Института пчеловодства РАСХН и его подразделений. Изучали пчел из популяций Татарстана, Башкирии, Марий Эл, Кемеровской, Вологодской, Кировской, Орловской областей, Мордовии, Красноярского и Пермского краев. По морфологическим характеристикам все они соответствовали критериям темной лесной пчелы. Исследование маркерной области генома митохондрий выявило наряду с уже известными фрагментами длиной 600 п. н., 800 п. н., характерными для A. mellifera mellifera, и тяжелые фрагменты длиной около 1200 п. н., первичная структура которых уточняется (рис. 2).
Авторы этой статьи в целях паспортизации использовали 24 семьи из коллекции Института пчеловодства РАСХН и его подразделений. Изучали пчел из популяций Татарстана, Башкирии, Марий Эл, Кемеровской, Вологодской, Кировской, Орловской областей, Мордовии, Красноярского и Пермского краев. По морфологическим характеристикам все они соответствовали критериям темной лесной пчелы. Исследование маркерной области генома митохондрий выявило наряду с уже известными фрагментами длиной 600 п. н., 800 п. н., характерными для A. mellifera mellifera, и тяжелые фрагменты длиной около 1200 п. н., первичная структура которых уточняется (рис. 2).
Обнаружена необычайно высокая концентрация тяжелых вариантов 800–1200 п. н. в большинстве исследованных популяций. Остается неизвестным, с чем связана столь высокая концентрация тяжелых фрагментов? Интересными представляются вопросы об адаптивной ценности таких маркеров, а также вопросы о механизмах их возникновения и об эволюционной истории подвида A. mellifera mellifera в России.
Таким образом, полученные нами результаты паспортизации свидетельствуют о сохранении среднерусской породы пчел в отдельных регионах средней полосы России, что имеет большое значение для охраны и интродукции ее генофонда.
Выполненное исследование — первый этап большой работы по разработке надежных критериев генетической паспортизации пород пчел России. В настоящее время она приобретает особое значение в связи с тем, что основным направлением современного промышленного пчеловодства являются создание и поддержание материнских и отцовских линий с последующими межлинейными скрещиваниями, с помощью инструментального осеменения и отбор эффективных комбинаций среди межлинейных гибридов.
Настоящее исследование генетической структуры популяций пчел Apis mellifera проводили в рамках проекта целевых исследований РФФИ «Разработка ДНК-технологий для паспортизации пород и породных линий медоносной пчелы Apis mellifera на территории России» (грант №08-04-13740-офи_ц).
М.А.МОНАХОВА,
И.И.ГОРЯЧЕВА,
Н.И.КРИВЦОВ
Московский государственный университет им. М.В.Ломоносова,
Институт общей генетики им. Н.И.Вавилова,
Научно-исследовательский институт пчеловодства РАСХН
Ключевые слова:
пчелы, паспортизация пород пчел, ДНК-маркер.
Аннотация:
сообщается о разработке критериев генетической паспортизации пород пчел России на основе ДНК-маркеров.
Summary:
it's informed on working out of criteria of genetic certification of breeds of bees of Russia on the basis of DNA-markers.
Keywords:
bees, species of bees, DNA marker.
Литература:
1. Ильясов Р.А., Поскряков А.В., Николенко А.Г. Методы идентификации подвида пчелы медоносной (A.m. mellifera L.). // Пчеловодство. — № 8. — 2008. — с. 8-9.
2. Монахова М.А. Генетические основы феномена пестрого расплода. // Пчеловодство. — № 1. — 2008. — с. 16-18.
- ВКонтакте
- РћРТвЂВВВВВВВВнокласснРСвЂВВВВВВВВРєРСвЂВВВВВВВВ




Автомобильное загрязнение и качество про…
нояб 10, 2016
Особенности пчеловодного сезона…
нояб 17, 2015
Акарасан при варроатозе и акарапидозе…
апр 11, 2015
Заслонка для летков
март 5, 2018
Английские впечатления…
фев 1, 2017
«Искры» между цветками и шмелями…
нояб 29, 2017
Биологические параметры особей пчелиных …
фев 25, 2023
Сырость губит пчел
окт 26, 2020
Методика определения пыльцы в меде…
янв 9, 2017









 Адрес редакции журнала "Пчеловодство":
Адрес редакции журнала "Пчеловодство":