Количество тепла, выделяемого семьей в единицу времени и фиксируемого ульем-калориметром, определяют соотношением
Количество тепла, выделяемого семьей в единицу времени и фиксируемого ульем-калориметром, определяют соотношением
Q = KF(tср.об – tн) = KFΔt,
где К — коэффициенты теплопередачи внутриульевой среды наружной, Вт/м2 . град; F — площадь поверхности ограждения, м2; tн — температура наружного воздуха, °С; tcp.oб — среднеобъемная температура в улье, °С;
tcp.oб=(Σ tiVi)/V,
где ti и Vi — температура, С°, и объем, м3, отдельных нагретых зон; V — внутренний объем улья, м3.
При постоянных величинах внешней температуры, объема, поверхности и коэффициента теплопередачи уровень термогенеза гнезда тем выше, чем выше температура нагретой зоны и чем на больший объем она распространяется. Вполне очевидно, как объем нагретой зоны, так и температура в ней, а также цель, с которой она создается, могут быть различными. Так, в летнее время в гнезде есть зоны расплода, переработки нектара, строительства сотов и т.п. Однако с какой бы целью их не создавали, температура в них никогда не превышает предельной 36°С, если даже назначения нагретых зон совпадают. Например, если в гнезде большой объем расплода, а температура в улье близка к предельной, то принос небольшого количества нектара или весенняя подкормка не вызовут дополнительного возрастания температуры и роста термогенеза.
В течение ряда лет мы проводили наблюдения за термогенезом семей в активный период, используя улей-калориметр. Следует заметить, что само начало активного сезона — это понятие условное. Для семей, находящихся в зимовнике или других помещениях, за начало активного сезона принимают день их выставки, а для зимующих на воле — день массового облета. Мы наблюдали его 4–10 апреля. При этом термогенез возрастал с 8–10 Вт (в конце зимовки) до 16–18 Вт и держался на этом уровне несколько дней, вплоть до раздачи побудительной подкормки и беглого осмотра. В данном случае речь идет о массовом лете пчел, хотя небольшой неспровоцированный их лет можно было наблюдать и раньше этого срока. Так, весной 2000 г. массовый вылет пчел приходился на 4 апреля, одиночные вылеты мы наблюдали с 24 марта, однако они не приводили к существенному росту термогенеза. После беглого осмотра и побудительной подкормки его уровень возрос в 2001 г. до 24 Вт. Через 10 дней он снизился и до 20 июля держался на уровне 20–22 Вт, отклоняясь в отдельные периоды в бóльшую сторону. Мы отмечали это 3–15 и 20–30 мая, когда термогенез в среднем составлял 25–26 Вт, и в период медосбора с 22 июня по 16 июля: термогенез 23 Вт. С конца июля и по 26 августа обсуждаемый показатель держался на уровне 12 Вт. Вмешательство пчеловода при формировании гнезда (26 августа) привело к повышению его уровня до 17 Вт (на 10 дней). До 20 Вт термогенез повысился при осенней подкормке (9–20 сентября) и сохранялся в последующий период на уровне 15 Вт, то есть на 3 Вт выше, чем до сборки гнезд и подкормки. При лечебно-профилактической обработке семьи бипином его показатель на протяжении 4 дней был на уровне 15 Вт.
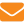
 Наши наблюдения за ряд лет подтвердили положения, представленные в формуле. Каждый раз, когда расширялась нагретая зона и повышалась ее температура, возрастал и термогенез. Так, например, во время медосбора с 1–11 июля 2000 г. он возрос с 22 до 30 Вт. При осенней подкормке 7–15 сентября — с 13 до 26 Вт. Однако этот показатель зависит не только от состояния семьи и выполняемой пчелами работы, но и в определяющей степени от температуры внешней среды. Приведем пример такой зависимости по результатам наблюдений в активный период 2002 г. (рис. 1). Ее можно представить тремя характерными кривыми. Кривая 1 определяет первый период с 24 апреля (среднесуточная температура над гнездом — 30,7°С) по 6 августа (когда температура последний раз поднялась до 30,5°С) при наличии большого количества расплода и среднесуточной температуре 10–28°С. Энергетические затраты семьи изменялись с 12 Вт (27°С) до 33 Вт (10°С) (разброс точек составляет ±5 Вт от среднего значения). Температура над гнездом за весь период в среднем составила 30–34°С. Момент наступления его четко ограничен.
Наши наблюдения за ряд лет подтвердили положения, представленные в формуле. Каждый раз, когда расширялась нагретая зона и повышалась ее температура, возрастал и термогенез. Так, например, во время медосбора с 1–11 июля 2000 г. он возрос с 22 до 30 Вт. При осенней подкормке 7–15 сентября — с 13 до 26 Вт. Однако этот показатель зависит не только от состояния семьи и выполняемой пчелами работы, но и в определяющей степени от температуры внешней среды. Приведем пример такой зависимости по результатам наблюдений в активный период 2002 г. (рис. 1). Ее можно представить тремя характерными кривыми. Кривая 1 определяет первый период с 24 апреля (среднесуточная температура над гнездом — 30,7°С) по 6 августа (когда температура последний раз поднялась до 30,5°С) при наличии большого количества расплода и среднесуточной температуре 10–28°С. Энергетические затраты семьи изменялись с 12 Вт (27°С) до 33 Вт (10°С) (разброс точек составляет ±5 Вт от среднего значения). Температура над гнездом за весь период в среднем составила 30–34°С. Момент наступления его четко ограничен.
Второй период отражает кривая 2, которая относится ко времени, когда в семье было небольшое количество расплода. Складывается впечатление, что пчелы держали его на таком уровне, чтобы без энергетических затрат суметь обогреть гнездо. Этот период в семье дискретен и характерен для ранней весны и осени. В нашем случае — до 24 апреля и с 7 августа до начала сентября при внешних среднесуточных температурах от 10 до 22°С. Энергетические затраты семьи здесь также изменялись в зависимости от наружной температуры, но на более низком уровне: с 8 Вт при среднесуточной температуре воздуха 22°С до 20 Вт при 10°С (разброс точек редко превышал 2–3 Вт от среднего значения). Температура над гнездом колебалась от 22 до 29°С. Конец этого периода и переход к третьему при температурах 20°С и выше размыт. Видимо, в этот период пчелы лучше заботятся об энергосбережении, а поддерживают его за счет более компактного обсиживания гнезда и не стремятся нарастить к зиме более значительную массу. Иначе кривую 2 они бы легко «перевели» в кривую 1, к чему подталкивает их отсутствие поступления нектара и пыльцы.
Третий период отражает кривая 3, когда расплода в гнезде мало или он отсутствует, что характерно для осени. Его начало можно условно отнести к первой декаде сентября. При изменении наружной температуры с 22 до 10°С тепловыделение составляет 5–7 Вт, и лишь при ее падении с 10 до 0°С оно доходит до 9 Вт. Такое явление характерно для бездеятельного периода, когда в природе не было медосбора. Если пчелам дать подкормку, как это сделали 9 сентября, их термогенез переходит в зону второго или даже первого периода. Температура над гнездом после подкормки поднимается с 20,9 до 34,1°С, а термогенез — с 5,5 до 25 Вт. Однако это продолжается недолго: уже 12 сентября температура снизилась до 26°С, а к утру 14 сентября опустилась до прежнего уровня — 21,0°С. Примерно к этому же периоду вернулся к своим показателям и термогенез.
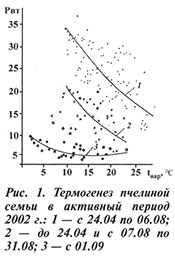
 Аналогичную картину динамики термогенеза в активный период прослеживали и в другие годы наблюдений, но с небольшими изменениями. Так, теплая погода в апреле и начале мая 2004 г. спровоцировала наращивание расплода, а последовавшая за этим холодная погода во второй половине мая привела к интенсивному росту термогенеза, связанному с его обогревом (кривая 1; рис. 2).
Аналогичную картину динамики термогенеза в активный период прослеживали и в другие годы наблюдений, но с небольшими изменениями. Так, теплая погода в апреле и начале мая 2004 г. спровоцировала наращивание расплода, а последовавшая за этим холодная погода во второй половине мая привела к интенсивному росту термогенеза, связанному с его обогревом (кривая 1; рис. 2).
Ранее мы наблюдали влияние похолодания на термогенез в конце апреля — начале мая, когда в семье было мало расплода. Пчелы, группируясь, легко закрывали его, и тепловыделение не только не возрастало, но и в некоторой степени падало. В рассматриваемом случае простой перегруппировкой обогреть расплод они не могли и вынуждены были прибегнуть к интенсификации теплообразования.
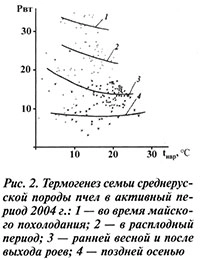
 В литературе много внимания уделяют искусственному обогреву гнезд в ранневесенний период. Многочисленные испытания этого приема не дают однозначного ответа на вопрос об его эффективности, в одних случаях эффект обогрева проявляется, в других — нет. Видимо, если похолодания были в начальный период, когда расплода мало, существенного эффекта от обогрева нет; когда же в гнезде много расплода, определенный эффект уже был: дополнительное выделение тепла в холодный период составляло в среднем 7,0–8,0 Вт. Семья в активный период дважды отроилась. Первый рой массой 3,5 кг вышел 19 июня, второй — массой 1,3 кг — 23 июня.
В литературе много внимания уделяют искусственному обогреву гнезд в ранневесенний период. Многочисленные испытания этого приема не дают однозначного ответа на вопрос об его эффективности, в одних случаях эффект обогрева проявляется, в других — нет. Видимо, если похолодания были в начальный период, когда расплода мало, существенного эффекта от обогрева нет; когда же в гнезде много расплода, определенный эффект уже был: дополнительное выделение тепла в холодный период составляло в среднем 7,0–8,0 Вт. Семья в активный период дважды отроилась. Первый рой массой 3,5 кг вышел 19 июня, второй — массой 1,3 кг — 23 июня.
Непосредственно перед роением ее теплообразование составляло 22–23 Вт. После выхода первого роя оно снизилось до 13(14) Вт, а после выхода второго — до 10(11) Вт (рис. 3). Рой вышел в 14 ч 15 мин, при этом до 14 ч никаких признаков возбуждения пчел не было, семья выделяла 16 Вт, затем в течение 15 мин она возбудилась, термогенез ее резко возрос, а после выхода роя она постепенно успокоилась, и через 45 мин выделение тепла остановилось на уровне 14 Вт. Выход столь большого количества пчел оказался губительным для семьи. Как видно из рисунка 2, ее термогенез упал до уровня ранневесеннего периода, оставаясь таким до конца сезона, и 3 октября подопытная семья погибла.
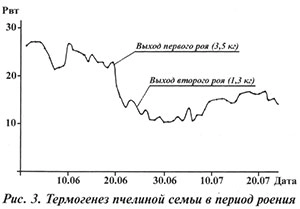
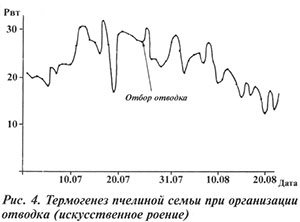 При искусственном роении организация отводка в 2003 г. существенно не повлияла на состояние семьи, она продолжала нормально работать и развиваться (см. рис. 4).
При искусственном роении организация отводка в 2003 г. существенно не повлияла на состояние семьи, она продолжала нормально работать и развиваться (см. рис. 4).
Наши наблюдения позволили сделать следующие выводы. ♦ Термогенез семьи в активный период выражается затратами тепла на поддержание определенного уровня температуры в объеме гнезда. ♦ Существуют три четко выраженных уровня термогенеза: с третьей декады апреля по первую декаду августа; с третьей декады апреля и со второй декады августа до начала сентября; с начала сентября до начала зимовки. ♦ В весенне-летнее время термогенез как в первом, так и во втором периоде с понижением наружной температуры возрастает. ♦ В осенний период с понижением наружной температуры до 10°С термогенез семьи падает, а при дальнейшем его понижении снова растет, и семья переходит в режим зимовки. ♦ При резком и продолжительном похолодании, если в семье много расплода, когда пчелы посредством простого агрегатирования не в состоянии согреть гнездо, они прибегают к дополнительному образованию тепла, составляющему 7–8 Вт. В результате общее тепловыделение семьи в этот период доходит до 35 Вт. Этот дополнительный уровень тепловыделений (7–8 Вт) может служить обоснованием для выбора мощности установки для искусственного обогрева.
В результате роения термогенез семьи снижается на 8–10 Вт и более. Семья по своим показателям практически переходит на ранневесеннюю стадию своего развития и до конца активного сезона не доходит до нормальной кондиции.
Характерно, что при контролируемой организации отводка столь негативных последствий не отмечено.
А.И.КАСЬЯНОВ
НИИ пчеловодства
- ВКонтакте
- РћРТвЂВВВВВВВВнокласснРСвЂВВВВВВВВРєРСвЂВВВВВВВВ

«Детские» дозы пчелиного яда …
дек 14, 2017
Ульи с уменьшенными рамками…
июнь 18, 2015
Эноциты осенне-зимних пчел при ядоотборе…
авг 13, 2015
Заслонка для летков
март 5, 2018
Пчеловоды, шмелеводы нужны России…
окт 28, 2015
Селекция среднерусских пчел в республике…
дек 1, 2016
Простая и удобная кормушка…
авг 7, 2018
Доработка пчеловодного инвентаря …
март 27, 2021
Меченые матки облегчают работу…
авг 28, 2017
Испытания феромонных препаратов в Кемер…
июль 1, 2014
Некоторые способы поимки роев…
март 17, 2016
Медоносный потенциал и численность семей…
дек 4, 2017
История красноуфимской популяции среднер…
апр 25, 2018
Как я очищаю воск
дек 9, 2017
«Апимил» — приманка для уховерток…
окт 29, 2015








 Адрес редакции журнала "Пчеловодство":
Адрес редакции журнала "Пчеловодство":