УДК 595.79
Смена генераций рабочих особей пчелиной семьи, отличающихся морфометрическими признаками, происходит в процессе их репродукции и элиминации. Динамика этих процессов существенно изменяется в последовательности генераций пчел, достигая максимума в весенне-летний и понижаясь до минимума в осенне-зимний период. Поэтому в годичном цикле жизни состав рабочих особей в семье характеризуется наибольшей изменчивостью морфометрических признаков в весенне-летний период.
Весной замене перезимовавших пчел чаще всего сопутствует уменьшение размеров крыльев. К середине лета они обычно увеличиваются. В дальнейшем (к концу лета — началу осени) с равной вероятностью возможно как увеличение, так и уменьшение размеров крыльев. Динамика их изменчивости в течение периода интенсивной репродукции пчел может существенно отклоняться от указанной тенденции даже в семьях, находящихся в одинаковых экологических условиях. Например, от майской к июльской генерации пчел в одной из семей длина крыла возрастала в среднем на 2,2%, в другой — на 4,3%. От июльской к сентябрьской — у пчел первой семьи происходило уменьшение длины крыльев на 1,1%, у второй — увеличение на 2,5%.
Элиминация зимующих пчел обладает некоторой избирательностью. Она выражается в том, что за время зимовки в наибольшей мере элиминируются пчелы с относительно небольшими крыльями [4]. В результате этого до начала весны доживают преимущественно пчелы с относительно большими крыльями. У весенних генераций пчел возможна противоположная тенденция изменчивости размеров крыльев. Например, элиминация части пчел от начала к окончанию зимы повлияла на увеличение в среднем длины передних крыльев у пчел, доживавших до весны на 0,7%, задних — на 0,8%. Весной за время смены перезимовавших пчел длина их крыльев в тех же семьях уменьшалась на 2,9 и 3,0% (P ≥ 0,99) соответственно. Это обусловливалось неблагоприятными для жизни пчелиных семей и развития пчел условиями (похолоданиями и невозможностью пополнения кормовых запасов).
Температура оказывает существенное влияние на развитие пчел. У рабочих особей, маток и трутней размеры крыльев различаются, но достигают максимума в случае развития при оптимальной температуре. Отклонение температуры от оптимальной оказывает неодинаковое влияние на уменьшение размеров крыльев. Если у рабочих особей, маток и трутней среднеевропейских пчел длина крыльев в случае развития при 34°С составляет в среднем (9,37±0,016), (10,08±0,026) и (12,41±0,031) мм, то при 30°С она уменьшается на 2,8; 2,1 и 3,1%, а при 36,5°С — на 1; 1,2 и 3,9% соответственно. Подобно этому изменяется и ширина крыльев.
Температура влияет на жилкование крыльев. Длина костальной жилки, как и всего крыла, достигает максимума у пчел, развивающихся в области температурного оптимума. При понижении температуры с 34 до 29,5°С длина жилки уменьшается в среднем на 4,5%, а при повышении с 34 до 37°С — на 0,9%. Изменения длины радиальной жилки в указанных отклонениях температуры от 34°С достигают 5,1 и 2% соответственно, а анальной — 0,3 и 5,6% соответственно.
Длина поперечных жилок в отличие от продольных (костальной, анальной и др.) уменьшается с повышением температуры инкубации от нижней к верхней границе витального диапазона. В частности, понижение температуры с 37 до 29,5°С уменьшает длину второй возвратной жилки на 9,2%, невральной — на 12,1%.
Вероятность возникновения аномалий жилкования возрастает с приближением температуры на стадиях предкуколки и куколки к границам витального диапазона (29...37°С). Понижение температуры до 29,5°С приводит к возникновению разрывов у жилки, разделяющей вторую и третью кубитальные ячейки, у (31±12)% пчел. Приобретение ответвления (нового отростка) у этой жилки отмечалось примерно у 2% пчел, развивавшихся у нижней температурной границы витального диапазона.
Значительные аномалии жилкования вызывают кратковременные воздействия гипо- или гипертермии. У (1±0,3)% пчел, охлаждавшихся на стадии яйца, или предкуколки, или куколки среднего возраста в течение (1,5±0,5) ч до 0°С, на стадии имаго обнаружено изменение конфигурации жилок и образование дополнительных отростков. Аномалии кубитальной, радиальной и дискоидальной ячеек обнаружены у (6,6±0,4)% пчел, которых на эмбриональной стадии подвергали гипертермии при 50°С в течение 45 мин.
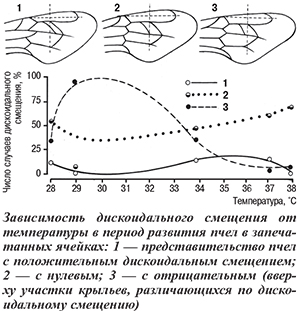 От температуры в период развития от предкуколки до имаго зависит дискоидальное смещение, которое рекомендуется использовать для дифференциации внутривидовых группировок (рас и популяций) медоносной пчелы [1]. Значение дискоидального смещения определяют по положению соединения задней межкубитальной жилки с радиальной и вершиной угла, образуемого возвратной и субдискоидальной жилками. Смещение этого угла к дистальной части крыла принято считать положительным, к его основанию — отрицательным, промежуточное — нулевым (рис.).
От температуры в период развития от предкуколки до имаго зависит дискоидальное смещение, которое рекомендуется использовать для дифференциации внутривидовых группировок (рас и популяций) медоносной пчелы [1]. Значение дискоидального смещения определяют по положению соединения задней межкубитальной жилки с радиальной и вершиной угла, образуемого возвратной и субдискоидальной жилками. Смещение этого угла к дистальной части крыла принято считать положительным, к его основанию — отрицательным, промежуточное — нулевым (рис.).
Представительство пчел с разным дискоидальным смещением варьирует в течение сезона их воспроизводства и может различаться в одной семье в одно и то же время. Причина этого вероятнее всего температура, которая различается на сотах, занятых расплодом [4]. В частности, на расплоде одних и тех же семей, инкубируемом при оптимальной температуре (34...35°С), примерно одинаковое представительство имели особи со всеми тремя формами смещений, при 29°С доминировало отрицательное смещение, и при 37...38°С на 25% понижалась доля положительного и отрицательного смещений, а нулевое — составляло около 50% (см. рис.).
Некоторое влияние оказывает температура на структуру аппарата (краевые складки и зацепки), сцепляющего передние крылья с задними. Длина краевых складок, посредством которых задние крылья сцепляются с передними, существенно уменьшается только в случае понижения температуры от оптимальной к нижней границе ее витального диапазона. Повышение температуры незначительно влияет на длину этих складок. У пчел, длина краевых складок которых при 34°С равнялась 2,49 мм, при понижении температуры до 29,5°С уменьшалась на 12,9%, а при увеличении до 37°С изменялась всего на 0,3%.
На задних крыльях размер участков, занимаемых зацепками, и их число зависят от температуры в период развития пчел в запечатанных ячейках: размер уменьшается в среднем на 6% при понижении температуры с 37 до 29°С, число зацепок — с 19,9 до 17,2 при том же диапазоне понижения температуры.
С изменчивостью размеров крыльев, их жилкованием и строением аппарата, сцепляющего крылья, связана их асимметричность. Вероятность симметричности крыльев возрастает с приближением их размеров к максимальным значениям, которые достигаются в случае развития в области оптимальных температур. Однако у некоторой части этих пчел возможно несовпадение по размерам крыльев. У пчел, развивающихся при 29°С, максимальное неравенство по длине левых и правых крыльев может достигать 2,9%, при 34°С — 1,9%, при 37°С — 2%, а по ширине — 2,8; 4,1 и 3,8% соответственно. Сходная несоразмерность свойственна задним крыльям. Несоответствие числа зацепок на левых и правых задних крыльях встречается примерно у трети пчел.
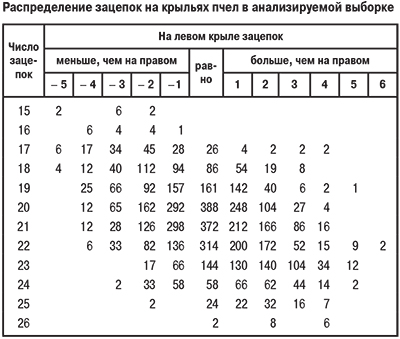 Посредством сопоставления строения аппаратов, сцепляющих передние и задние крылья, у каждой из отдельно взятых пчел (табл.) впервые была выявлена закономерность изменчивости асимметричности крыльев по этому признаку [2]. Установлено, что вероятность асимметричности правых и левых передних крыльев зависит от длины краевых складок, а задних — от числа зацепок. Соответственно закономерность изменчивости симметричности крыльев выражается в переориентации доминирования по числу зацепок с право- на левостороннее в зависимости от увеличения их числа (см. табл.). Аналогичную зависимость имеет связь асимметричности крыльев от их размеров и длины краевых складок [2]. Очевидно, в этом выражается общая закономерность связи асимметричности с размерами парных органов, что подтверждается исследованиями на бумажных осах [3, 5] и позволяет проследить использование распределения по признакам каждой особи в анализируемой выборке (см. табл.).
Посредством сопоставления строения аппаратов, сцепляющих передние и задние крылья, у каждой из отдельно взятых пчел (табл.) впервые была выявлена закономерность изменчивости асимметричности крыльев по этому признаку [2]. Установлено, что вероятность асимметричности правых и левых передних крыльев зависит от длины краевых складок, а задних — от числа зацепок. Соответственно закономерность изменчивости симметричности крыльев выражается в переориентации доминирования по числу зацепок с право- на левостороннее в зависимости от увеличения их числа (см. табл.). Аналогичную зависимость имеет связь асимметричности крыльев от их размеров и длины краевых складок [2]. Очевидно, в этом выражается общая закономерность связи асимметричности с размерами парных органов, что подтверждается исследованиями на бумажных осах [3, 5] и позволяет проследить использование распределения по признакам каждой особи в анализируемой выборке (см. табл.).
В течение имагинальной стадии пчел от их младшего к старшему возрасту происходит избирательная элиминация по размерам крыльев. При прочих равных условиях в наибольшей мере элиминируются обладатели крайних вариантов по этому признаку. В результате диапазон изменчивости их размеров сокращается на 19–28%. Соответственно увеличивается представительство пчел с высокой выраженностью симметричности. Из этого следует, что асимметричность размеров крыльев и их сцепляющих аппаратов, приобретаемая некоторой частью пчел к началу имагинальной стадии, не имеет для них адаптивного значения.
Таким образом, изменчивость строения, симметричности и размеров крыльев зависит от условий развития пчел до стадии имаго: в оптимальных условиях снижается вероятность возникновения аномалий жилкования крыльев. В течение жизни на стадии имаго элиминируются преимущественно особи, у которых размеры крыльев отличаются от средней нормы. Симметричность крыльев по размерам и строению сцепляющих аппаратов у пчел возрастает с приближением этих морфометрических параметров к средней норме. Зависимость дискоидального смещения от температуры исключает использование этого признака для внутривидовой дифференциации медоносной пчелы.
Е.К.ЕСЬКОВ, М.Д.ЕСЬКОВА
ФГБОУ ВО «Российский государственный
аграрный заочный университет»
Изменчивость строения, симметричности и размеров крыльев зависит от условий развития пчел в период, предшествующий стадии имаго. При развитии пчел в оптимальных условиях снижается вероятность возникновения аномалий жилкования крыльев. В течение жизни на стадии имаго элиминируются преимущественно особи, у которых размеры крыльев отличаются от средней нормы. Симметричность крыльев по размерам и строению сцепляющих аппаратов у пчел возрастает с их приближением по этим морфометрическим признакам к средней норме. Зависимость дискоидального смещения от температуры исключает использование этого признака для внутривидовой дифференциации медоносной пчелы.
Ключевые слова: крылья, симметричность, жилкование, аномалии, изменчивость.
ЛИТЕРАТУРА
1. Билаш Г.Д., Кривцов Н.И. Селекция пчел. — М., 1991.
2. Еськов Е.К. Зависимость асимметричности аппаратов, сцепляющих крылья пчел, от вариабельности количества краевых зацепок // Докл. РАН. — 1997. — Т. 352. — № 4.
3. Еськов Е.К., Еськова М.Д., Спасик С.Е., Тобоев В.А. Соразмерность и асимметричность крыльев у шершня обыкновенного (Hymenoptera, Vespinae) // Евроазиатский энтомологический журнал. — 2015. — Т. 14. — № 1.
4. Еськов Е.К. Эволюция, экология и этология медоносной пчелы. — М., 2016.
5. Еськов Е.К., Еськова М.Д., Тобоев В.А. Закономерности изменчивости асимметричности ячеек сот и крыльев у медоносной пчелы и бумажных ос // Успехи современной биологии. — 2017. — Т. 138. — № 2.
СВЕДЕНИЯ ОБ АВТОРАХ:
Еськов Евгений Константинович, д-р биол. наук, проф., заслуженный деятель науки и техники РФ, e-mail:
Еськова Майя Дмитриевна, д-р биол. наук, проф.
VARIATION OF SIZE, VENATION AND SYMMETRY OF THE WING OF THE HONEY BEE
E.K.Eskov, M.D.Eskova
The variability of the structure, symmetry and wing size depends on conditions of bee development during the period preceding the imago stage. Likelihood of wing venation anomalies decreases when bees develop in optimal conditions. During the life at the imago stage individuals that have different dimensions of the wings from the average rate are primarily eliminated. The wings symmetry on the size and structure of the bees hamuli increases at the approach of these morphometric characteristics to the average rate. The disk-like offset dependence on the temperature excludes the use of this feature for intraspecific honey bee differentiation.
Keywords: symmetry, venation, abnormalities variability.

Опыт бессотовой зимовки…
окт 15, 2014
Семью подсиливаю роями …
сен 1, 2015
Пчелы готовятся к зимовке…
сен 18, 2014
Вы решили приобрести пакет пчел……
июнь 1, 2024
Экспертиза качества меда…
нояб 5, 2015
Периоды в годовом цикле жизни пчел…
июнь 20, 2024
Майские пакеты
апр 18, 2015
Павильонные летковые заградители…
сен 26, 2014
Породная характеристика пчел…
окт 4, 2022
Степные шмели
нояб 21, 2014
Динамика изменения экстерьерных признако…
апр 7, 2015
Некоторые советы по содержанию пчел в ул…
авг 6, 2016
Электрообогрев (5)
фев 18, 2016
Весеннее наращивание пчел…
мая 27, 2022
Основные болезни медоносных пчел в зимне…
сен 23, 2017








 Адрес редакции журнала "Пчеловодство":
Адрес редакции журнала "Пчеловодство":