


 Еще задолго до К.Линнея англичанин Рэй (J.Ray) в 1710 г. назвал медоносную пчелу, разводимую на крестьянских подворьях, Apis domestica — пчела домашняя. Разумеется, это было неверно, поскольку и до сих пор пчелу медоносную назвать домашней в полном смысле этого слова нельзя. К.Линней, давший научное название виду пчелы медоносной Apis mellifera, имел в виду прежде всего местную шведскую пчелу и совсем не подозревал о многообразии внутривидовых форм. Поздние исследователи выделили мелкие таксономические единицы (подвиды, расы, экотипы), в значительной мере отличающиеся друг от друга комплексом хозяйственно полезных и биологических признаков, сформировавшихся в различающихся природно-климатических условиях и адаптированные к ним в процессе длительной селекции.
Еще задолго до К.Линнея англичанин Рэй (J.Ray) в 1710 г. назвал медоносную пчелу, разводимую на крестьянских подворьях, Apis domestica — пчела домашняя. Разумеется, это было неверно, поскольку и до сих пор пчелу медоносную назвать домашней в полном смысле этого слова нельзя. К.Линней, давший научное название виду пчелы медоносной Apis mellifera, имел в виду прежде всего местную шведскую пчелу и совсем не подозревал о многообразии внутривидовых форм. Поздние исследователи выделили мелкие таксономические единицы (подвиды, расы, экотипы), в значительной мере отличающиеся друг от друга комплексом хозяйственно полезных и биологических признаков, сформировавшихся в различающихся природно-климатических условиях и адаптированные к ним в процессе длительной селекции.
По классификации F.Ruttner (1992) на земном шаре насчитывается 25 подвидов (рас, пород) пчелы медоносной. Кроме хорошо всем известных европейских пород — темной лесной (A.m.mellifera), серой горной кавказской (A.m.caucasica), желтой долинной кавказской (A.m.remipes), итальянской (A.m.ligustica), краинской (A.m.carnica), карпатской (A.m.carpatica) и украинской степной (A.m.acervorum) следует назвать сирийскую пчелу (A.m.syriaca), палестинскую (A.m.saneta), анатолийскую серую (A.m.insularium), иранскую (A.m.iranica), североафриканскую черную (A.m.intermissa), рифскую (A.m.major), сахарскую (A.m.sahariensis), египетскую (A.m.lamarckii), суданскую (A.m.nubica), восточно-африканскую (A.m.scutellata), горную (A.m.monticola), западно-африканскую (A.m.adansonii), капскую (A.m.capensis), мадагаскарскую (A.m.unicolor), йменскую (A.m.jemenitica). Разумеется, это далеко не полный перечень. Следует учесть, что внутри подвидов сложились различные популяции и экотипы. В связи с этим со всей остротой вырисовывается проблема их идентификации. Более того, в процессе селекционной работы создаются новые таксоны (линии, внутрипородные типы) и их также необходимо идентифицировать.
Классический метод различения внутривидовых таксонов — морфометрический анализ. Его основу заложил Г.А.Кожевников (1900, 1906). Он еще в 1898 г. сделал первые промеры длины хоботка кавказских пчел и предложил методику измерения их хитиновых частей.
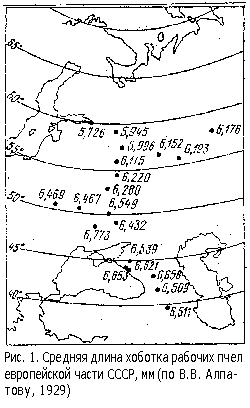 А.С.Михайлов (1924), W.W.Alpatov (1925) ее усовершенствовали и установили зависимость размеров хитиновых частей пчел (прежде всего длины хоботка) от географической широты местности (рис.1). Это давало возможность идентифицировать отдельные таксоны из географических мест, удаленных по долготе. Однако метод не срабатывал при оценке таксонов, обитающих на одной и той же широте. Кроме того, морфологические признаки (прежде всего длина хоботка) подвержены сезонным изменениям, которые в ряде случаев превышают географические различия. В этом случае становится перспективным сравнивать изменчивость экстерьерных признаков: абсолютных значений коэффициентов вариации (Cv, %) и среднеквадратичных отклонений (d).
А.С.Михайлов (1924), W.W.Alpatov (1925) ее усовершенствовали и установили зависимость размеров хитиновых частей пчел (прежде всего длины хоботка) от географической широты местности (рис.1). Это давало возможность идентифицировать отдельные таксоны из географических мест, удаленных по долготе. Однако метод не срабатывал при оценке таксонов, обитающих на одной и той же широте. Кроме того, морфологические признаки (прежде всего длина хоботка) подвержены сезонным изменениям, которые в ряде случаев превышают географические различия. В этом случае становится перспективным сравнивать изменчивость экстерьерных признаков: абсолютных значений коэффициентов вариации (Cv, %) и среднеквадратичных отклонений (d).
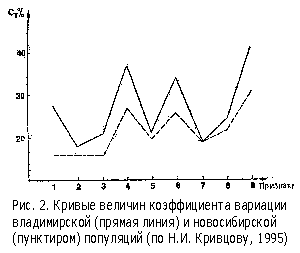
Возможность использования параметров изменчивости для изучения популяционной структуры вида на разных зоологических объектах показали А.В.Яблоков (1966), А.П.Разоренова (1975), Ю.И.Новоженов (1970). С помощью наших исследований мы подтвердили перспективность этого метода (рис. 2) для идентификации различных популяций среднерусской породы пчел (Н.И.Кривцов, 1995).
 Оценочную шкалу идентификации пчел по окраске, а также по дискоидальному смещению в жилковании крыла разработал G.Goetze (1930, 1940, 1949, 1964). Необходимо отметить, что некоторые подвиды пчел различаются по окраске хитина, и этот признак пригоден для их определения, но его можно использовать лишь в первом приближении.
Оценочную шкалу идентификации пчел по окраске, а также по дискоидальному смещению в жилковании крыла разработал G.Goetze (1930, 1940, 1949, 1964). Необходимо отметить, что некоторые подвиды пчел различаются по окраске хитина, и этот признак пригоден для их определения, но его можно использовать лишь в первом приближении.
Хорошим дополнением и даже альтернативой морфологическим методам идентификации внутривидовых таксонов может быть оценка поведенческих или этологических реакций пчел. В настоящее время используется в той или иной степени сравнение таких поведенческих признаков как злобивость, ройливость, гигиенические особенности поведения, мобилизационная активность на источники медосбора, флоромиграция и флороспециализация, поведение на соте при осмотре гнезда, прополисование гнезда, печатка меда и т.д. Очевидно, может быть перспективным изучение адаптационных приспособлений к условиям среды (характер развития, реакция на неблагоприятные условия и т.д.). Однако большинство этих признаков качественные и методики их оценки в большинстве случаев несовершенны, а полученные разными авторами данные в силу вышеуказанных причин противоречивы, а посему и несопоставимы. Вместе с тем Е.К.Еськов (1966) показал, что акустический сигнал дальности является породным признаком пчел. И.А.Левченко (1969) отметил, что в активности сигнальной деятельности и характере танца имеются межпородные различия.
В плане дифференциации мелких таксонов следует обратить внимание и на некоторые физиологические признаки пчел. Так, М.В.Жеребкин (1979) показал существенные различия активности каталазы ректальных желез пчел среднерусской и кавказской пород, отмечаются различия и в активности других ферментов (диастаза, инвертаза) у разных пород пчел (М.В.Жеребкин, В.П.Чаплыгин, 1969).
На цитологические и химические особенности, присущие разным породам пчел, обратил внимание М.Г.Стелвизов (1963, 1976). Это разница в размерах клеточных ядер, содержании ДНК в ядрах соматических тканей.
M.Bounias (1976, 1978) нашел несоответствия в белковых спектрах гемолимфы рабочих пчел различных пород. Он отметил разницу содержания в показателях трегалозы, глюкозы и фруктозы в гемолимфе этих насекомых.
В последние годы делаются попытки применения метода ДНК-маркирования, и прежде всего для генетической паспортизации многих видов сельскохозяйственных животных при селекционной работе. У пчел относительно хорошо изучены митохондриальные ДНК-маркеры, но из-за матроклинного характера наследования они не дают возможность анализировать межпородные гибриды. В то же время показано, что RAPD-технологии при правильном подборе параметров и отборе соответствующих праймеров вполне пригодны для проведения эффективной геномной дактилоскопии различных подвидов (пород) пчел (В.Н.Поздняков, 2000; О.С.Чудинов, 2002 и др.). Эти методики дают возможность отслеживать различное происхождение пчел на основании исследования геномной ДНК.
 Метод молекулярно-генетического контроля ДНК и ДНК-маркирования перспективен для идентификации пород и результатов селекционных достижений и, очевидно, займет в недалеком будущем лидирующее положение (рис. 3).
Метод молекулярно-генетического контроля ДНК и ДНК-маркирования перспективен для идентификации пород и результатов селекционных достижений и, очевидно, займет в недалеком будущем лидирующее положение (рис. 3).
Поскольку морфолого-географический метод обладает ограниченной различительной способностью, с его помощью можно улавливать лишь географические расы (подвиды). Только комплексные генетико-эколого-географические исследования позволяют наиболее полно вскрыть структуру вида.
К.М.Завадский (1968) предлагает следующую внутривидовую структуру: полувид — раса, почти достигшая состояния молодого вида; подвид — сформировавшаяся географическая или экологическая раса; экотип — локальная экологическая раса; местная популяция — занимает определенную территорию и способна к самовоспроизведению; экоэлемент — внутрипопуляционная форма; морфобиологическая группа — группа внутри популяции; биотип — это элементарная единица генетической структуры популяции. В состав популяций входят формы, проверенные отбором, но любая селекционная работа нарушает ее историческую структуру, искажает предмет исследования (С.А.Северцев, 1951), поэтому такие новообразования требуют объективной идентификации.
В пчеловодстве, как уже было сказано выше, в лучшем случае выделяются экотипы (Ж.Луво, 1969), поэтому для тонкого изучения структуры вида пчелы медоносной (Apis mellifera) предстоит большая и кропотливая работа систематического плана, базирующаяся на солидной методологической базе.
Н.И.КРИВЦОВ
























 Адрес редакции журнала "Пчеловодство":
Адрес редакции журнала "Пчеловодство": +7 (499) 270-05-59
+7 (499) 270-05-59


