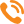Полет пчел обеспечивает асинхронная мускулатура — наиболее совершенный вид мускулатуры беспозвоночных. Функциональная единица здесь — многоядерное мышечное волокно, образующееся при эмбриональном развитии в результате слияния отдельных клеток. В основе его сокращения лежат конформационные изменения внутриклеточных белковых комплексов — актина и миозина.
Полет пчел обеспечивает асинхронная мускулатура — наиболее совершенный вид мускулатуры беспозвоночных. Функциональная единица здесь — многоядерное мышечное волокно, образующееся при эмбриональном развитии в результате слияния отдельных клеток. В основе его сокращения лежат конформационные изменения внутриклеточных белковых комплексов — актина и миозина.
В результате их взаимодействия происходит гидролиз молекул АТФ, и химическая энергия превращается в механическую работу мышечного сокращения. С точки зрения интенсивности работы (количества используемой АТФ в единицу времени) асинхронные летательные мышцы насекомых представляют собой наиболее рациональную механохимическую сократительную систему.
Для изучения особенностей структурной организации мышечных волокон в онтогенезе мы провели исследование летательной мышцы медоносной пчелы методами световой и электронной микроскопии. Кусочки мышечной ткани объемом 1 мм3, а также отдельные изолированные мышечные волокна фиксировали в 1%-ном растворе ОsО4 на веронал-ацетатном буфере (рН 7,4) по Колфилду и после стандартной проводки через спирты и ацетон заливали в аралдит или эпон-аралдит.
Готовили полутонкие и ультратонкие срезы, контрастировали нитратом свинца и просматривали в электронном микроскопе JЕМ-100. Мерные показатели органелл (число в единице объема цитоплазмы, объемные доли, площади поверхностей) определяли методами Э.Р.Вейбеля (1970) и Р.К.Данилова и В.Н.Сперанского (1988) непосредственно на негативах, увеличенных на увеличителе «Беларусь» при Ку = 3,5. Активность АТФ-азы определяли методом Падикула и Герман при рН 9,4; сукцинатдегидрогеназы (СДГ) — тетразолиевым методом Нахласа.
 По результатам электронно-микроскопических исследований мы можем представить следующие данные (рис.1). На рисунке 1 представлены электроннограммы продольного среза летательной мускулатуры 30-суточных рабочих пчел. Видны структурные компоненты мышечного волокна. Темные Z-диски делят мышечное волокно на отдельные саркомеры. В середине саркомера заметна узкая темная М-полоска. На электроннограмме видны отличительные особенности асинхронных мышечных волокон: I диски практически не просматриваются, все пространство саркомера занято А диском. Между миофибриллами располагаются плотноприлегающие крупные митохондрии. Отсутствует саркоплазматическая сеть. Видны прилегающие к митохондриям просветы трахеол. На стадии куколки площадь мышечного волокна равна 79,0 мкм2; диаметр миозиновых миофиламентов — 122 А° ; число мышечных волокон в мышце — 145 шт. Число митохондрий в симпластах куколки в суммарном объеме составляет 14% от общего объема клетки. При этом доля митохондрий, расположенных по периферии клетки, в 2,13 раза превышает долю интерфибриллярных митохондрий.
По результатам электронно-микроскопических исследований мы можем представить следующие данные (рис.1). На рисунке 1 представлены электроннограммы продольного среза летательной мускулатуры 30-суточных рабочих пчел. Видны структурные компоненты мышечного волокна. Темные Z-диски делят мышечное волокно на отдельные саркомеры. В середине саркомера заметна узкая темная М-полоска. На электроннограмме видны отличительные особенности асинхронных мышечных волокон: I диски практически не просматриваются, все пространство саркомера занято А диском. Между миофибриллами располагаются плотноприлегающие крупные митохондрии. Отсутствует саркоплазматическая сеть. Видны прилегающие к митохондриям просветы трахеол. На стадии куколки площадь мышечного волокна равна 79,0 мкм2; диаметр миозиновых миофиламентов — 122 А° ; число мышечных волокон в мышце — 145 шт. Число митохондрий в симпластах куколки в суммарном объеме составляет 14% от общего объема клетки. При этом доля митохондрий, расположенных по периферии клетки, в 2,13 раза превышает долю интерфибриллярных митохондрий.
У пчел 1–3-суточного возраста мерные показатели достоверно увеличены: площадь мышечных волокон — до 92,0 мкм2; диаметр миозиновых миофиламентов — до 140 А° ; число мышечных волокон в мышце — до 185 шт. Число интерфибриллярных митохондрий в мышечных волокнах становится равным числу периферических. Суммарное их число хотя и превышает показатель стадии куколки, однако остается еще невысоким — 25% от общего объема клетки. Уровень окислительно-восстановительных процессов на кристах митохондрий невысок, о чем свидетельствует низкая активность (СДГ) — 0,18 усл. ед. На этой стадии зарегистрирован минимальный для имаго показатель гистохимической активности миозиновой и митохондриальной АТФ-аз, ответственных за расщепление макроэргических связей в молекуле АТФ и обеспечение энергией процессов мышечного сокращения — 1,12 и 0,52 усл. ед. соответственно.
Морфометрические и гистохимические показатели летательной мышцы рабочих пчел 4–7-суточного возраста свидетельствуют о значительной физиологической перестройке: площадь мышечного волокна составляет 106 мкм2; диаметр миозиновых миофиламентов — 175 А° ; число мышечных волокн в мышце — до 226 шт., что превышает соответствующие показатели на стадии куколки в 1,34; 1,43 и 1,22 раза. Возрастает до 36% доля митохондрий в объеме симпласта продольной грудной мышцы. Соотношение периферических и интерфибриллярных митохондрий меняется в сторону преобладания последних (1:2,52). Активность СДГ увеличивается в 13,5 раза по сравнению с предыдущей группой и достигает 2,43 усл. ед. Гистохимическая активность АТФ-аз миозина и митохондрий регистрируется на уровне 3,3 и 2,33 усл. ед. Достигнутый на этой стадии уровень обменных процессов уже достаточен для кратковременного полета, так как именно в этом возрасте мы регистрировали вылет меченых пчел на первые очистительные и ориентировочные облеты.
В возрасте 8–10 сут площадь мышечных волокон по сравнению с показателями куколки увеличивается в 1,39 раза и составляет 110 мкм2. Диаметр миозиновых нитей возрастает до 186 А° ; число мышечных волокн в мышце — до 228 шт. Общее число митохондрий возрастает настолько, что они занимают 55% объема мышечного волокна. Число интерфибриллярных митохондрий превышает число периферических в 3,45 раза. Активность СДГ в 18,94 раза выше, чем в первой имагинальной группе, и составляет 3,41 усл. ед.
У пчел 11–20-суточного возраста наблюдается значительное повышение метаболической активности летательной мускулатуры. Хотя общее число митохондрий увеличивается незначительно (до 58%), а соотношение интерфибриллярных и периферических несколько снижается, активность окислительно-восстановительных процессов на кристах митохондрий резко возрастает: уровень СДГ повышается до 8,19 усл. ед. (превышает уровень активности этого фермента у 1–3-суточных пчел в 45,5 раза). Активность АТФ-азы миозина достигает 6,52 усл. ед., митохондриальной АТФ-азы — 15,24 усл. ед., то есть превышают показатели первой имагинальной стадии соответственно в 5,82 и 29,3 раза.
 Подготовка летательной мускулатуры к функционированию во время длительного полета проявляется и положительной динамикой морфометрических показателей структурных единиц мышечного волокна и нарастанием активности ферментов, обеспечивающих энергией процессы сокращения. Площадь мышечных волокон по сравнению с куколкой увеличивается в 1,46 раза и составляет 115 мкм2; диаметр миозиновых нитей возрастает до 222 А° ; число мышечных волокон в мышце в этот период несколько снижается (213 шт.), однако разница с предыдущей группой недостоверна (Р < 0,95). В этом возрасте пчелы начинают вылетать для сбора нектара, и в нашем опыте к летной деятельности приступили 80% меченых пчел.
Подготовка летательной мускулатуры к функционированию во время длительного полета проявляется и положительной динамикой морфометрических показателей структурных единиц мышечного волокна и нарастанием активности ферментов, обеспечивающих энергией процессы сокращения. Площадь мышечных волокон по сравнению с куколкой увеличивается в 1,46 раза и составляет 115 мкм2; диаметр миозиновых нитей возрастает до 222 А° ; число мышечных волокон в мышце в этот период несколько снижается (213 шт.), однако разница с предыдущей группой недостоверна (Р < 0,95). В этом возрасте пчелы начинают вылетать для сбора нектара, и в нашем опыте к летной деятельности приступили 80% меченых пчел.
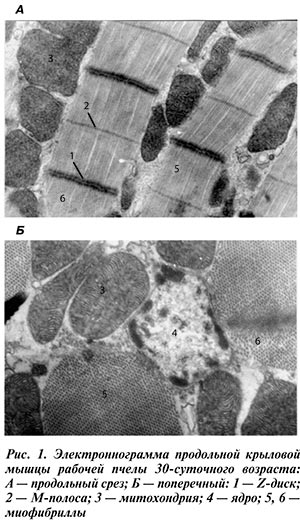
После 20-го дня жизни при активной собирательной деятельности морфометрические показатели летательной мышцы стабилизируются: площадь мышечного волокна — на уровне 114 мкм2; диаметр миозиновых миофиламентов — 221 А° ; число мышечных волокон в мышце — на уровне 229 шт. Заметно возрастает активность миозиновой АТФ-азы (9,35 усл. ед.), а активность митохондриальной остается практически на прежнем уровне — 15,31 усл. ед. Период жизни пчел с 21-го по 30-й день характеризуется активной летной деятельностью и требует максимальных затрат энергии, и мы отмечаем еще большее увеличение разницы между числом интерфибриллярных и периферических митохондрий. Их соотношение в симпласте составляет 3,78:1. Активность СДГ повышается по сравнению с первой имагинальной стадией в 48 раз и составляет 8,65 усл. ед. (рис. 2).
Таким образом, в летательной мускулатуре пчел в процессе миогистогенеза происходят генетически детерминированные закономерные качественные и количественные перестройки структурных компонентов мышечного волокна — контрактильного и энергетического аппаратов. В постэмбриональном гистогенезе диаметр миозионовых миофиламентов, представляющих контрактильный аппарат, увеличивается в 1,8 раза, а объем энергетического аппарата (суммарный объем митохондрий) — на 44%. Терминальная дифференцировка мышечных волокон при подготовке рабочих пчел к активной летной деятельности ориентирована на выполнение длительной сократительной функции мускулов. Площадь мышечного волокна увеличивается с 79,0 мкм2 на стадии куколки до 115 мкм2 (в 1,45 раза) у летных рабочих пчел; существенно повышается роль интерфибриллярных митохондрий, обеспечивающих высокую ферментативную активность в дефинитивных мышечных волокнах и адаптацию мышечной ткани к возрастанию нагрузки.
Г.С.МИШУКОВСКАЯ*, А.Г.МАННАПОВ**
* Башкирский ГАУ
** РГАУ – МСХА им. К.А.Тимирязева, г. Москва

Кто поселился в тростнике?…
июль 13, 2015
Защита пчел от повышенной влажности во в…
дек 15, 2020
И от роения может быть польза…
июль 14, 2016
Оригинальная маточная клеточка…
июль 17, 2015
Использование апипродуктов в офтальмолог…
окт 24, 2014
Связанная вода в теле пчел и зимостойкос…
дек 4, 2015
Особенности применения препарата «АПИРОЙ…
мая 18, 2015
Отделение меда от воска при распечатыван…
фев 13, 2015
Залог здоровья пчел
июнь 1, 2022
Пчеловодство в двухматочных ульях Озеров…
сен 30, 2014
Безроевая система с тихой сменой матки п…
июнь 19, 2014
Образование зимнего клуба в малоформатны…
дек 1, 2021
Успешное лечение пчел
июль 6, 2017
Загадки медоносных растений…
дек 2, 2015
Ветеринарная санитария на пасеках…
июнь 28, 2023







 Адрес редакции журнала "Пчеловодство":
Адрес редакции журнала "Пчеловодство":