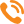Генетический состав естественных популяций пчел неоднороден. Он характеризуется большим набором разнокачественных генов.
Генетический состав естественных популяций пчел неоднороден. Он характеризуется большим набором разнокачественных генов.
Баланс генов в популяциях каждой породы исторически складывался под воздействием естественного отбора. В результате появились адаптивные гены с комбинаторными особенностями, позволяющие популяциям приспособиться к условиям среды обитания.
Естественно сложившаяся популяция обладает присущей ей изменчивостью признаков и свойств, характеризующейся нормой реакции, означающей, что каждый признак может изменяться в границах генетически определенных для него значений. Норма реакции зависит от многих причин внутреннего характера и условий внешней среды.
Совокупность генов популяции называется генофондом. Каждая порода в условиях ее естественного обитания обладает своим исторически сложившимся уникальным генофондом со свойственным ему динамическим равновесием, которое обеспечивается механизмами передачи наследственного материала.
 На схеме (рис. 1) показана генетическая взаимосвязь при передаче наследственного материала в популяции пчел.
На схеме (рис. 1) показана генетическая взаимосвязь при передаче наследственного материала в популяции пчел.
Центральное звено этой связи — трутни. Они осуществляют передачу геномов от матки-матери матке-дочери.
При замене матки в каждой семье, выведенной в ней же маткой-дочерью, генофонд популяции и его состояние сохраняются.
Матки-дочери не наследуют всех признаков материнской семьи (матка-дочь получает через трутня геном другой матки), и их свойства зависят от сочетания генов их геномов с генами полученного генома.
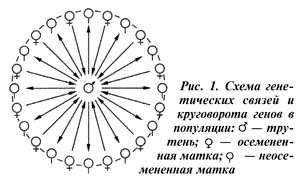
 Хозяйственно ценные качества семьи зависят не только от матки, но и от ее дочерей — рабочих пчел. При этом генотип матки не совпадает с генотипом ее потомства, так как генотип рабочих пчел образуется из генома матки-матери и геномов других маток, переданных трутнями (рис. 2).
Хозяйственно ценные качества семьи зависят не только от матки, но и от ее дочерей — рабочих пчел. При этом генотип матки не совпадает с генотипом ее потомства, так как генотип рабочих пчел образуется из генома матки-матери и геномов других маток, переданных трутнями (рис. 2).
Вследствие того, что матка осеменена многими трутнями, происходящими от других маток, генотипы рабочих пчел будут различны.
В популяции пчел происходит довольно сложный и уникальный процесс передачи наследственного материала.
 Кроме плановой замены маток пчеловод ведет непрерывный отбор. Необходимость его объясняется проявлением изменчивости в популяции.
Кроме плановой замены маток пчеловод ведет непрерывный отбор. Необходимость его объясняется проявлением изменчивости в популяции.
Как правило, 15–20% семей по уровню продуктивности ниже среднего показателя для популяции.
Слабые семьи он присоединяет к другим семьям, уничтожая маток, а часть подсиливает расплодом от других семей. И то и другое дает только временный эффект повышения продуктивности. Такие матки подлежат замене.
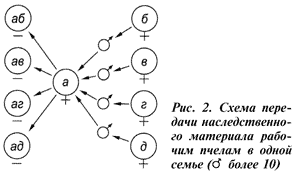
Казалось бы, что отбором, особенно интенсивным, мы нарушаем генный баланс пасеки в сторону увеличения числа более продуктивных семей в следующих поколениях. Однако этого не происходит, и в популяции восстанавливается генное равновесие в прежних пределах. Такое свойство популяции получило название популяционный генетический гомеостаз (рис. 3).
В.П.НИКОЛАЕНКО
г. Ростов-на-Дону

Многокорпусный улей перспективен…
мая 26, 2020
Весы для контроля за медосбором…
март 24, 2015
Распространение аскосфероза в Узбекистан…
март 27, 2017
Сачок для снятия роев
сен 5, 2021
Эноциты осенне-зимних пчел при ядоотборе…
авг 13, 2015
Влияние стимулирующих подкормок на прием…
апр 29, 2023
Почему гибнут пчелы
дек 22, 2023
Революция или эволюция?…
март 8, 2015
Какие ульи были в имении Пушкина?…
июнь 11, 2018
Нектароносные липняки…
сен 23, 2015
Пищевая ценность сахара и меда…
фев 1, 2015
Купание пчелы
июль 21, 2017
Объединение пчелиных семей осенью (2)…
сен 1, 2019
Мед в медицине
июль 9, 2016
Подагра, артрозы, остеохондроз…
апр 3, 2018








 Адрес редакции журнала "Пчеловодство":
Адрес редакции журнала "Пчеловодство":