УДК 595.799:638.1
В организации целенаправленного поведения пчел важная роль принадлежит химической ориентации. Она включает выделение и восприятие ими собственных пахучих веществ, а также запахов врагов, природных и синтетических веществ. Реагирование на химические раздражители обеспечивается функционированием дистантных (действующих на расстоянии) и контактных (действующих при прикосновении) хеморецепторов. Качество и пригодность для употребления трофических субстратов контролируются вкусовыми рецепторами.
У рабочих пчел, маток и трутней в секреции пахучих веществ (феромонов) участвуют специализированные органы, локализующиеся в разных частях тела. В 1882 г. Н.В.Насонов обнаружил у рабочих пчел тергальные железы, получившие название «Насоновы». Позже такие железы были найдены и у маток.
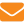
Насоновы железы представлены скоплением крупных субэпидермальных клеток, протоки которых проходят через тергит или межсегментную мембрану. У маток железы располагаются у оснований второго, третьего и четвертого тергитов, у рабочих пчел — между шестым и седьмым тергитом (рис. 1).
 Феромон активно функционирующей Насоновой железы содержит 1,8 мкг гераниола, 1,4 мкг нероловой кислоты, 0,8 мкг фарнезола, 0,2 мкг гераниоловой кислоты, а также цис- и транс-изомеры цитраля [8]. Судя по возрастной динамике изменения состава феромона, интенсивное функционирование железы приходится на период активной летной деятельности пчелы. У 15-дневных особей железа еще не выделяет гераниол, процесс начинается с 20-го дня жизни при уровне основного компонента секрета 1,2 мкг [3].
Феромон активно функционирующей Насоновой железы содержит 1,8 мкг гераниола, 1,4 мкг нероловой кислоты, 0,8 мкг фарнезола, 0,2 мкг гераниоловой кислоты, а также цис- и транс-изомеры цитраля [8]. Судя по возрастной динамике изменения состава феромона, интенсивное функционирование железы приходится на период активной летной деятельности пчелы. У 15-дневных особей железа еще не выделяет гераниол, процесс начинается с 20-го дня жизни при уровне основного компонента секрета 1,2 мкг [3].
Секреторная активность тергальных желез маток достигает максимума в 1–2-недельном возрасте, то есть в возрасте половой зрелости и готовности к спариванию. Секрет обладает аттрактивностью для трутней [4], и они используют его для опознания маток, влетающих в их скопление.
Рабочие пчелы для распространения запаха секрета Насоновой железы выпячивают ее, поднимая вверх брюшко (см. рис. 1). В такой позе они нередко задерживаются у входа в жилище и машут крыльями, чем усиливают распространение феромона. Пчелы-фуражиры выпячивают железу при повторных посещениях кормушек во время наполнения зобиков растворами углеводов высокой концентрации. Но это несвойственно пчелам-фуражирам вида Apis mellifera, посещающим цветки медоносных растений, хотя типично для близкого ему вида Apis indica [4].
Выпячивание Насоновой железы пчелами-фуражирами начинается во время полета к неоднократно посещаемой кормушке [6]. Если она находится на небольшом расстоянии от гнезда, то большинство пчел, перемещающихся по опорному субстрату, выпячивают железу, когда направляются к кормушке. При этом имеет значение преодолеваемое расстояние. Среди пчел, посещавших кормушку в 0,1 м от летка, всего 3,3% выпячивали железу, а в 1, 3 и 5 м — соответственно 22, 68 и 84%. При расположении кормушки в 10 м от летка около 80% пчел выпячивают железу только вблизи кормушки [2].
Время, необходимое пчеле для заполнения медового зобика, обычно немного меньше продолжительности периода, в течение которого она выпячивает железу. На заполнение зобика 50%-ным раствором сахарозы пчела затрачивает (71,8±1,9) с, а железа закрывается через (58,3±4,1) с. Время, в течение которого пчела выпячивает железу, находясь на кормушке, сокращается соответственно уменьшению количества забираемого корма и его концентрации. Ароматизация корма или пространства вокруг кормушки уменьшает число пчел, выпячивающих железу. Однако существенное значение имеет плотность пчел на кормушке: на площади 1 или 20 см2 выпячивание железы прекращается, когда на ней собирается соответственно 4–7 или 20–25 особей.
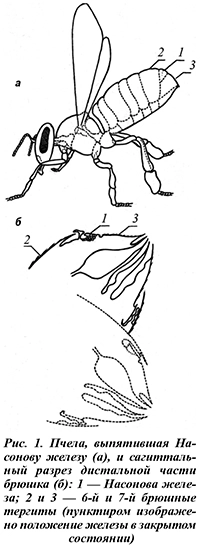
В консолидации пчелиной семьи важная роль принадлежит секреторной функции мандибулярных желез матки, хотя они имеются также и у рабочих пчел, и у трутней. Железы располагаются в голове над основанием мандибул. Секрет выделяется через протоки, заканчивающиеся у основания мандибул. У матки железы отличаются наибольшим размером, у трутней — наименьшим. При прочих равных условиях размер железы зависит от возраста и физиологического состояния насекомого. От начала имагинальной стадии к третьему дню жизни размер железы у матки увеличивается примерно в 1,5 раза. У рабочих пчел мандибулярные железы достигают максимальных размеров к началу имагинальной стадии при условии, что на стадии куколки поддерживалась стабильная температура на уровне 34°С. Их продольная ось составляет 1,22 мм, поперечная — 0,62 мм (рис. 2), что в среднем на 2% больше, чем у пчел, развивавшихся при 30 или 37°С [1].
 У маток прослеживается прямая связь между активностью репродуктивной функции и количеством транс-9-оксодецен-2-овой кислоты, секретируемой мандибулярными железами. У плодных маток в 3-недельном возрасте мандибулярные железы содержат около 47 мкг феромона, к 4-недельному возрасту его количество возрастает до 240 мкг, к 3-месячному — до 360 мкг, а в возрасте 2–3 лет снижается до 70 мкг [1].
У маток прослеживается прямая связь между активностью репродуктивной функции и количеством транс-9-оксодецен-2-овой кислоты, секретируемой мандибулярными железами. У плодных маток в 3-недельном возрасте мандибулярные железы содержат около 47 мкг феромона, к 4-недельному возрасту его количество возрастает до 240 мкг, к 3-месячному — до 360 мкг, а в возрасте 2–3 лет снижается до 70 мкг [1].
Для восприятия растворенных в воздухе различных химических веществ у пчел, как и у многих других видов насекомых, имеются специализированные сенсиллы. Они различаются по строению и локализации. Хеморецепторный антеннальный комплекс (сенсиллы, расположенные на антеннах) обеспечивает восприятие химических веществ, распространяющихся по воздуху (дистантная хеморецепция), а также посредством контакта с различными субстратами (контактная хеморецепция). При этом сенсиллы, относящиеся к одному и тому же типу, могут выполнять разные функции. Это связано со строением их кутикулярных аппаратов и функциональной организацией. Контактная трихоидная сенсилла отличается наличием толстостенного волоска с одной или несколькими порами, локализованными в его дистальной части. Поры могут закрываться кутикулярными клапанами. Для дистантных трихоидных сенсилл характерно наличие тонкостенных волосков, пронизанных большим числом постоянно открытых пор.
На антеннах рабочей пчелы и трутня локализуется около 98% дистантных рецепторов. В опытах с последовательной ампутацией члеников антенн у пчел, повторно прилетавших к кормушке с ароматизированным кормом, установлено, что удаление 2–3 члеников снижало число случаев правильного выбора кормушки на 5–6%, 4–6 члеников — на 8–11%, 7 члеников — на 24%. Потеря способности находить кормушку с ароматизированным кормом происходила при удалении всех 8 члеников антенн [5].
В поведенческих опытах у пчел, подлетающих к кормушкам с известным запахом корма, определены пороги чувствительности. За пороговые значения пахучих веществ принимали такие их концентрации, которые привлекали фуражиров к кормушкам с испытуемым пахучим веществом. Установлено, что пчелы обнаруживают запахи гераниола и метилгептинола при растворении 1 г этих веществ в 1•109 г этилгликогеля. Чувствительность к запахам фенилпропилалкоголя, фенилпропилацетата и ряда жирных кислот резко падает после снижения их концентрации с 1,9•109 до 4,3•1011 моль/см3. С 95,5%-ной вероятностью пчелы в состоянии различить 1816 парных сочетаний запахов [11].
Чувствительность хеморецепторов пчелы и человека к одним пахучим веществам в значительной мере совпадает, к другим — существенно различается. Пчелы превосходят человека по чувствительности к запахам цветков многих энтомофильных растений. К этиленгликолю они чувствительнее человека примерно в 40 раз, а к метилгептенону — в 100 раз [9]. Но чувствительность человека к эвганолу превосходит чувствительность пчел более чем в 200 раз, к скатолу — примерно в 800 раз. Особенно пчелы восприимчивы к феромонам маток.
Электрофизиологические исследования показали высокую чувствительность плакоидных сенсилл рабочих пчел и трутней к экстрактам маток: 1 мкг этого вещества достаточно для изменения частоты пульсации рецептора. Реакция сенсилл антеннального комплекса на один из главных компонентов феромона маток — транс-9-оксодецен-2-овую кислоту — регистрируется при концентрации ее на уровне 108 молекул/см3 [11].
Реагирование пчел на экстракт матки не обладает видовой специфичностью. Рабочие особи медоносной пчелы адекватно реагируют как на экстракт маток своего вида, так и на экстракт маток индийской пчелы. Подобно этому, индийские пчелы не различают феромоны маток вида A. indica и A. mellifera. В исследованиях на индийской и медоносной пчелах не обнаружено различий по аттрактивности маток для трутней того или другого вида [10].
Пчел отличает от многих других животных и человека наличие специализированного рецептора СО2, роль которого выполняют сенсиллы ампуловидного и целоконического типов. Другая особенность этих рецепторов связана с их высокой чувствительностью к изменению температуры. Спонтанная импульсация их составляет около 5 Гц. При повышении СО2 в воздушной среде на 0,5% частота пульсаций рецептора в первые 0,2 с достигает 35 Гц, на 2% — 45 Гц, на 5% — 55 Гц и на 100% — 135 Гц. В процессе адаптации рецептора частота генерируемых им пульсаций через 1 с при указанных значениях СО2 понижается до 10–15, 20, 30 и 45 Гц [7].
Реагирование пчел на изменение влажности воздуха указывает на наличие у них соответствующих рецепторов. На антеннах насекомых обнаружены рецепторы, избирательно реагирующие на повышение или понижение влажности воздуха. Роль этих рецепторов выполняют целоконические, ампуловидные и целосферические сенсиллы. Механизм их действия, по-видимому, связан со смещениями, происходящими в кутикулярном аппарате под действием изменения температуры и/или различного насыщения воздуха водяными парами. Локализация этих сенсилл в кутикулярных углублениях экзоскелета антенн обеспечивает им защиту от механического воздействия при прикосновении антенн к различным субстратам.
При понижении влажности с 64 до 5% частота высокоамплитудных импульсов возрастает до 45 Гц, постепенно урежаясь к концу первой секунды до 30 Гц. За это время частота низкоамплитудных импульсов понижается с 12 до 6 Гц. В отличие от этого при повышении влажности с 5 до 64% частота высокоамплитудных импульсов понижается примерно с 35 до 17 Гц, а низкоамплитудных — возрастает с 10 до 45 Гц.
Итак, химическая ориентация пчел основана на использовании собственных пахучих выделений и запаха объектов внешней среды. Запах, ассоциируемый с кормом, используется пчелами-фуражирами в качестве локального ориентира. В отличие от этого феромоны, выделяемые пчелами у обильных источников корма, обладают аттрактивностью для других пчел-фуражиров. Это способствует снижению затрат энергии пчел на трофическое обеспечение пчелиных семей. Рецепторы, реагирующие на изменение содержания в воздухе водяных паров и диоксида углерода, используются пчелами для контроля и регуляции внутригнездового микроклимата.
Е.К.ЕСЬКОВ, М.Д.ЕСЬКОВА
Рассмотрены химическая сигнализация и пространственная ориентация пчел, основанная на использовании собственных пахучих выделений и запахов объектов внешней среды. Показано, что запах, ассоциируемый с кормом, используется пчелами-фуражирами в качестве локального ориентира. В отличие от этого феромоны, выделяемые пчелами у обильных источников корма, обладают аттрактивностью для других пчел-фуражиров. Рецепторы, реагирующие на изменение содержания в воздухе водяных паров и диоксида углерода, используются пчелами для контроля и регуляции внутригнездового микроклимата.
Ключевые слова: экзосекреторные железы, хеморецепторы, феромоны, химическая ориентация.
ЛИТЕРАТУРА
1. Еськов Е.К. Эволюция, экология и этология медоносной пчелы. — М., 2016.
2. Левченко И.А., Шалимов И.И. Железа Насонова у пчел-сборщиц // Пчеловодство. — 1975. — № 9.
3. Boch R., Shearer D.A. Production of geranial by honey bees of various ages // J. Ins. Physiol. — 1963. — V. 9. — № 4.
4. Butler C.G. Pheromones of queen honeybees // Rep. Pothamsted Exp. Sta. — 1969, 1970. — Part I.
5. Dostal B. Riechfahigkeit und Zalf der Riechsinneselements bei der Honigbiene // Zeit. vergl. Physiol. — 1958. — Bd. 41. — № 2.
6. Frisch K. von. Tanzsprache and Orientirung der Bienen. Berlin, Heidelberg, New-York: Springer-Verlag. 1965.
7. Lacher V. Electrophysiologische Untersuchungen an einzeinen Receptoren fur Geruch, Kohlendioxud, Luftfeuchtigkeit und Temperatur auf Antennen der Arbeitsbiene und der Drohne // Zeit. verge. Physiol. — 1964. — Bd. 48. — № 6.
8. Pickett J.A., Williams I.H., Martin A.P. et. al. Nasonov pheromone of the honey bee, Apis mellifera L. (Hymenoptera: Apidae) Part I. Chemical characterization // J. Chem. Scol. — 1980. — V. 6. — № 2.
9. Ribbands C.R. What can honey-bee smell? // Austral. Beekeeping. — 1956. — V. 57.
10. Ruttner F., Kaissling K.E. Uber die interspezifische Wirkung des Sexuallockstoffes von Apis mellifera und Apis cerana // Zool vergl. Physiol. — 1968. — Bd. 59.
11. Vareschi E. Duftuterscheidung bei der Honigbiene-Einzelzelt-Ableitungen und Verhaltensreaktionnen // Zeit. vergl. Physiol. — 1971. — Bd. 75. — № 2.
СВЕДЕНИЯ ОБ АВТОРАХ:
Еськов Евгений Константинович, д-р биол. наук, проф., заслуженный деятель науки и техники РФ, e-mail:
Еськова Майя Дмитриевна, д-р биол. наук, проф.
BEE SECRETION AND PERCEPTION OF ODOROUS SUBSTANCES
E.K.Eskov, M.D.Eskova
The paper studies the chemical signalizing and the spatial orientation of bees, based on the use of its own odorous emissions and odors of objects in the external environment. It is shown that smell associated with food, is used by the bee-foragers as a local landmark. In contrast, the pheromones emitted by the bees at rich sources of feed, have the attractiveness to other bee-foragers. Bees use receptors, respondent to changes in the air, water vapor and carbon dioxide, to control and regulate microclimate in a beehive.
Keywords: exosecretory glands, chemoreceptors, pheromones, chemical orientation.
- ВКонтакте
- РћРТвЂВВВВВВВВнокласснРСвЂВВВВВВВВРєРСвЂВВВВВВВВ

Как лучше использовать рои…
март 18, 2014
Техника против вирусов?…
янв 22, 2014
Комплексное оздоровление часто болеющих …
дек 12, 2017
Чудо-воскотопка
фев 14, 2015
Продукты пчел в лечении трофических язв…
авг 17, 2016
Биоэнергетика в пчеловодстве…
дек 30, 2015
Перерабатываем восковое сырье (1)…
дек 3, 2014
Геоактивные структуры земли и жизнедеяте…
фев 20, 2015
Аномалии жилкования крыльев трутней серо…
нояб 18, 2016
Сахар и уксус
фев 13, 2017
Распечатывание сотов
апр 25, 2016
Профилактика и лечение депрессии…
март 31, 2021
Термогенез семьи в активный период…
янв 27, 2021
Причина коллапса пчелиных семей…
сен 10, 2015
Работа пчел-фуражиров на Кубани …
март 18, 2015








 Адрес редакции журнала "Пчеловодство":
Адрес редакции журнала "Пчеловодство":