Явление пестрого расплода хорошо знакомо пчеловодам. Оно связано с ранней гибелью части личинок, в результате чего после запечатывания расплода часть ячеек остается пустой или повторно засеянной, с личинками разного возраста. Число пустых ячеек значительно варьирует и в отдельных семьях может достигать 50%.
Явление пестрого расплода хорошо знакомо пчеловодам. Оно связано с ранней гибелью части личинок, в результате чего после запечатывания расплода часть ячеек остается пустой или повторно засеянной, с личинками разного возраста. Число пустых ячеек значительно варьирует и в отдельных семьях может достигать 50%.
В этом случае обычно винят матку (старая, больная, слабая) и стремятся ее заменить на другую, чаще со своей же пасеки, но все опять повторяется, поскольку истинная природа этого явления связана с особенностями генетических механизмов определения пола у пчелы медоносной.
Впервые мысль о гапло-диплоидном механизме определения пола у пчелы была высказана Дзержоном более 150 лет тому назад (Dzierzon, 1861): матка и рабочие пчелы развиваются из оплодотворенных, а трутни — из неоплодотворенных яиц.
Почти через столетие О.Маккензен (О.Mackensen, 1951), изучая процесс наследования при близкородственных скрещиваниях (скрещивание матки с одним из братьев — трутнем), обнаружил явление пестрого расплода и дал ему генетическое обоснование. Он сравнивал полученные результаты с результатами наследования у наездника-хабробракона семейства браконид (отряд перепончатокрылых), у которого кроме гапло-диплоидного механизма определения пола был обнаружен ген пола «Х». В популяции он представлен серией множественных аллелей (различные состояния одного и того же гена). При записи аллелей этого гена используют буквенное обозначение: Ха, Хb, Хс, Хd и т.д.
Маккензен предположил, что аналогичный механизм определения пола может быть и у пчел. Из оплодотворенных яиц развиваются нормальные особи женского пола только в случае гетерозиготного состояния аллелей, то есть ХаХb, ХаХс, ХbХс и др. Гомозиготные же по аллелям этого гена зародыши (ХаХа, ХbХb, ХсХс и др.) развиваются в диплоидных трутней. Гомозиготное состояние аллелей гена пола имеет полулетальный эффект, такие зародыши обладают пониженной жизнеспособностью и погибают на ранних стадиях развития, давая картины пестрого расплода.
Предположение Маккензена было блестяще экспериментально подтверждено работами Войке (1963) с помощью искусственного осеменения пчелиных маток трутнями-братьями. Эти результаты были доложены на XIX Международном конгрессе по пчеловодству.
Оригинальность данной работы заключается в том, что автор определял пол насекомого еще в запечатанном расплоде по анатомическим особенностям зачатков половых органов личинок. Изучался расплод только тех маток, в семьях которых отмечалось около 50% пестрого расплода. Было обнаружено, что половина зародышей потомства таких маток несет признаки женского пола, а другая половина — мужского. Таким путем удалось доказать, что эмбриональная гибель зародыша связана с геном пола.
Автором было также высказано предположение о том, что диплоидные зародыши трутней погибают не сами по себе: их распознают и уничтожают рабочие пчелы сразу после выхода из яиц.
Почти одновременно с этой работой Лейдлов с соавторами (Laidlow et al., 1956), изучая распространение полулетальных генов в бразильской популяции более 60 пчелиных семей, обнаружили 12 аллелей этого гена.
Эволюция генетических механизмов определения пола у пчел. Таким образом у пчелы медоносной описано сосуществование двух систем определения пола: гапло-диплоидной и множественных аллелей гена пола. Предполагается, что это обстоятельство отражает эволюционные преобразования генетической системы определения пола, связанные с переходом пчелы от одиночного к семейному существованию (Шаскольский, 1971). Согласно автору, можно предположить существование трех этапов в становлении (совершенствовании) генетических механизмов систем определения пола у пчелы. Эти системы эволюционировали в связи с развитием и усложнением пчелиной семьи.
I этап. Исходная система — это самый древний механизм становления пола, определяется гетерохромосами (ХУ или ХО). Он широко распространен в мире насекомых. Такая система обеспечивает равновероятное появление особей мужского и женского пола в соотношении 1:1 при диплоидности самцов и самок.
Замена первой системы на вторую связана с прекращением одиночного существования и образованием семьи, специализацией особей в ней (матка, рабочие пчелы и трутни), резким сокращением числа размножающихся самок, уменьшение потребности в самцах сопровождалось совершенствованием генетических механизмов определения пола.
II этап. Система множественных аллелей гена пола. Пол определяется серией множественных аллелей гена пола. В этом случае самцов в семье рождается гораздо меньше (не половина всех особей), а только 2–8%, так как их появление определяется гомозиготным состоянием аллелей гена пола (ХаХа, ХbХb и др.). На этом этапе еще нет различий в плоидности, самки и самцы диплоидны.
III этап. Гапло-диплоидная система определения пола. Появление этой системы связано, по-видимому, с дальнейшим развитием семьи, увеличением численности ее особей до нескольких тысяч, при этом потребность в трутнях ограничивается только сезонным размножением.
Инстинкт удаления ненужных трутней сохраняется и в другом варианте: когда рабочие особи при резком похолодании или сокращении взятка выбрасывают уже зрелый трутневый расплод.
Гапло-диплоидный механизм регуляции пола позволяет пчелиной семье довольно легко регулировать время вывода трутней, а также их численность. Однако наряду с этим механизмом механизм множественных аллелей гена пола сохраняется в виде рудиментарной системы, которая заявляет о себе при близкородственном разведении феноменом пестрого расплода.
Геном трутня как генетический полигон. Переход на гапло-диплоидный механизм определения пола дал огромные преимущества медоносной пчеле в смысле цитогенетического механизма адаптации и ознаменовал исключительную роль трутней в этом процессе.
Гаплодная генетическая система самцов, обладающая одинарным набором хромосом, а следовательно и генов, представляет собой полигон, на котором выявляются все вредные рецессивные гены, снижающие жизнеспособность и адаптационный потенциал вида. Их проявление приводит к гибели особей, носителей этих генов. Таким образом, популяция имеет возможность освободиться от так называемого «генетического груза». Именно науровне генетической системы трутней идет отбор наиболее приспособленных к конкретным условиям существования генетических комбинаций. Поэтому, уничтожая трутневый расплод, мы тем самым препятствуем реализации одной из самых совершенных программ адаптации вида, которая совершенствовалась миллионы лет эволюции. Именно она придает генетическому аппарату пчелы удивительную пластичность, которая позволила Apis mellifera освоить все континенты, за исключением Антарктиды.
Интересно отметить, что гапло-диплоидный механизм определения пола широко распространен среди насекомых (более чем у 200 тыс. видов).
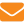
Множественные аллели гена пола на изолированной пасеке. Как уже отмечалось выше, большой процент пестрого расплода наблюдается именно на изолированных пасеках, где процветает близкородственное разведение. 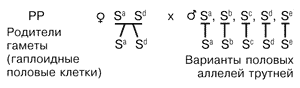 Проиллюстрируем это на схеме, используя обозначения аллелей гена пола Х. В зарубежной литературе их называют Sex-гены и обозначают символом S. Чтобы не путать Х — символ половых генов с Х-хромосомой, целесообразно использовать символ S для обозначения половых генов и в отечественных публикациях.
Проиллюстрируем это на схеме, используя обозначения аллелей гена пола Х. В зарубежной литературе их называют Sex-гены и обозначают символом S. Чтобы не путать Х — символ половых генов с Х-хромосомой, целесообразно использовать символ S для обозначения половых генов и в отечественных публикациях.
Рассмотрим на примере возможность комбинирования половых аллелей в локальной популяции.
В данном примере матка образует два типа гамет, каждая из которых несет по одному аллелю гена пола Sа или Sd. Гаплоидные трутни, а также их гаметы несут только по одному аллелю из возможной серии множественных аллелей: Sа или Sb, Sc, Sd и др. При оплодотворении возможны различные комбинации аллелей в зиготе. Для их воспроизведения воспользуемся решеткой, где по горизонтали расположим аллели гамет трутней, а по вертикали — матки. Для удобства используем только буквенные обозначения аллелей.
 Все гетерозиготные по гену пола особи будут нормальными особями женского пола, а гомозиготные — диплоидными трутнями, которых пчелы уничтожат, что и приведет к картине пестрого расплода. Если трутни окажутся братьями матки, несущими одинаковые с ней половые аллели, то процент пестрого расплода будет максимальным — 50%.
Все гетерозиготные по гену пола особи будут нормальными особями женского пола, а гомозиготные — диплоидными трутнями, которых пчелы уничтожат, что и приведет к картине пестрого расплода. Если трутни окажутся братьями матки, несущими одинаковые с ней половые аллели, то процент пестрого расплода будет максимальным — 50%.
Очевидно, чтобы избежать высокой концентрации одноименных аллелей на пасеке при искусственном выводе маток, необходимо использовать несколько донорских семей. Это особенно важно на изолированных пасеках.
В большинстве случаев пчелы сами преодолевают эту проблему. Случные пункты нередко бывают удалены от пасеки на значительные расстояния, что уменьшает вероятность родственного спаривания. С другой стороны, полиандрия (осеменение несколькими трутнями), несомненно, ценное биологическое приспособление, уменьшающее вероятность концентрации одноименных аллелей в одной популяции.
Проблема идентификации половых аллелей. Идентификация половых аллелей приобретает большое значение при искусственном осеменении маток. В этом случае генетический контроль производителей — важное условие селекционно-генетических исследований.
К сожалению, различные половые аллели не имеют фенотипического выражения, позволяющего проводить их идентификацию по внешнему виду. В связи с этим большое значение приобретает поиск их молекулярных маркеров. В настоящее время ведутся работы по картированию генома пчелы с их использованием. Уже получены первые результаты по картированию Sex-локуса (Hasselman et. al., 2004). Обнаружены фланкирующие последовательности, позволяющие идентифицировать половой локус и маркировать его на генетической карте (Beye et al., 1999). Одна из его отличительных особенностей — чрезвычайно высокий уровень рекомбинации, позволяющий определить его как «горячую точку». Обнаружен также большой полиморфизм фланкирующих участков по степени повторяемости. Все это свидетельствует о большом функциональном значении этого локуса в системе жизнеобеспечения вида. Sex-локус становится предметом изучения не только прикладной, но и фундаментальной науки.
М.А.МОНАХОВА
Секция «Биотехнологические
основы пчеловодства» МОИП
- ВКонтакте
- РћРТвЂВВВВВВВВнокласснРСвЂВВВВВВВВРєРСвЂВВВВВВВВ

Загадки в поведении пчел…
нояб 18, 2023
Ловите рои
июнь 4, 2016
Дезинфицирующее средство — препарат «Ве…
апр 1, 2015
Свечи на основе продуктов пчел в урологи…
сен 3, 2022
Медоносные растения периода динозавров…
окт 18, 2022
Помогла сырость
март 20, 2017
Селекция устойчивых к заболеваниям пчел …
мая 3, 2015
Как задержать рои
июль 9, 2015
Отбор медовых рамок и удаление пчел из м…
июнь 23, 2014
Доход в любой год
апр 8, 2015
Гибель пчел в странах Cеверной Америки и…
апр 8, 2019
Активность фосфатазы среднерусской и кар…
нояб 22, 2016
Развитие особей пчелиной семьи с учетом …
дек 11, 2015
Сетчатое ведро для облета пчел…
фев 15, 2015
Двухкорпусные семьи на слабом медосборе…
нояб 27, 2023








 Адрес редакции журнала "Пчеловодство":
Адрес редакции журнала "Пчеловодство":