Наследование большинства хозяйственных признаков пчел (как, впрочем, и у других сельскохозяйственных животных) обусловлено многими генами, каждый из которых оказывает лишь незначительное воздействие. При этом фенотипическое проявление полигенного признака в значительной степени зависит от преобладающих условий среды.
Наследование большинства хозяйственных признаков пчел (как, впрочем, и у других сельскохозяйственных животных) обусловлено многими генами, каждый из которых оказывает лишь незначительное воздействие. При этом фенотипическое проявление полигенного признака в значительной степени зависит от преобладающих условий среды.
Генетические параметры полигенных признаков определяются коэффициентами наследуемости, повторяемости и корреляции, которые целесообразно использовать при планировании селекционного процесса в пчеловодстве. Они рассчитываются генетико-статистическими методами (популяционная генетика возникла на основе синтеза генетики и математической статистики).
Наследуемость представляет собой долю участия наследственной (генотипической) изменчивости в общей фенотипической изменчивости признака конкретной популяции. Отношение генотипической вариансы к общей фенотипической Райт обозначил как коэффициент наследуемости h2. Чем выше значение h2 , тем больше влияние генотипической составляющей на проявление селекционного признака, а следовательно, больше возможностей его селекционного улучшения. Коэффициент наследуемости характеризует лишь ту долю наследственной изменчивости, которая обусловлена действием аддитивных факторов (неаллельных генов с внешне идентичным и кумулятивным действием).
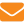
 Несмотря на определенные ограничения, показатель наследуемости широко используют в селекционной работе. Он позволяет судить о генотипическом разнообразии признака и дает возможность прогноза селекции по его фенотипическому проявлению уже на первых этапах работы. В таблице 1 приведены обобщенные данные разных авторов о значениях h2признаков пчел. Хотя сопоставление этих показателей и не совсем корректно по причине разных методов расчета, разных пород пчел и условий, все же анализ показывает, что коэффициенты наследуемости признаков, характеризующих продуктивность и плодовитость пчел, имеют значения на уровне таковых для традиционных в животноводстве объектов селекции.
Несмотря на определенные ограничения, показатель наследуемости широко используют в селекционной работе. Он позволяет судить о генотипическом разнообразии признака и дает возможность прогноза селекции по его фенотипическому проявлению уже на первых этапах работы. В таблице 1 приведены обобщенные данные разных авторов о значениях h2признаков пчел. Хотя сопоставление этих показателей и не совсем корректно по причине разных методов расчета, разных пород пчел и условий, все же анализ показывает, что коэффициенты наследуемости признаков, характеризующих продуктивность и плодовитость пчел, имеют значения на уровне таковых для традиционных в животноводстве объектов селекции.
Все приведенные в таблице 1 признаки имеют определенные перспективы в смысле селекции.
Особенно интересны данные по таким признакам, как реакция на феромон тревоги (при отборе на незлобливость) и гигиеническое поведение (при отборе на санирующие способности по очистке гнезда), так как по ним до сих пор не вели систематическую селекцию. Во всяком случае, в России пока нет линий пчел, отличающихся выраженным гигиеническим поведением, как и линий, отличающихся устойчивым миролюбием.
Работа в этих направлениях представляется очень перспективной. В отношении агрессивности С.Sport (1975) показал, что этот признак, относящийся к полигенным, частично доминирует в F1.
Повторяемость характеризует способность организмов удерживать свои показатели на том или ином уровне при изменении паратипических факторов (возраст, условия кормления, содержания и внешней среды и др.). Другими словами, речь идет о реализации наследственной информации в пределах одного поколения. По определению Н.А.Плохинского (1969), повторяемость возникает при реализации одной и той же групповой генетической информации у одних и тех же объектов в разном возрасте и при разных условиях. Так, лучшие в раннем возрасте не всегда остаются лучшими в последующем, лучшие в одних условиях жизни могут быть средними или даже худшими при других.
 Способность сохранять определенное постоянство признака имеет большое значение в селекционной работе. В определенном смысле повторяемость признака — высший предел его наследуемости. Коэффициент повторяемости (r) рассчитывают, как коэффициент корреляции по смежным учетам, показателям, измерениям. Чем больше таких корреляционных пар, тем надежнее показатели повторяемости. Более информативен квадрат коэффициента повторяемости (r2), дающий при умножении на 100 процентное выражение степени повторяемости. Для примера приводим наши данные (табл. 2) в отношении яйценоскости пчелиных маток.
Способность сохранять определенное постоянство признака имеет большое значение в селекционной работе. В определенном смысле повторяемость признака — высший предел его наследуемости. Коэффициент повторяемости (r) рассчитывают, как коэффициент корреляции по смежным учетам, показателям, измерениям. Чем больше таких корреляционных пар, тем надежнее показатели повторяемости. Более информативен квадрат коэффициента повторяемости (r2), дающий при умножении на 100 процентное выражение степени повторяемости. Для примера приводим наши данные (табл. 2) в отношении яйценоскости пчелиных маток.
Как видно, яйценоскость маток — достаточно устойчивый признак, доля действия факторов, общих для учетов в разном возрасте матки, проводимых при разных внешних условиях, составила в среднем 37%. Это обстоятельство открывает благоприятные перспективы для селекции на плодовитость маток.
 Подобный анализ целесообразно проводить по каждому селекционному признаку. Так, мы подтвердили, что среднерусские пчелы при прочих равных условиях лучше посещают гречиху, чем пчелы других пород (Д.В.Гирник, Н.И.Кривцов, 1989). Дальнейшие исследования показали, что признак «гречишности» отличается достаточно высокой повторяемостью (табл. 3). Доля действия общих факторов, определяющих проявление «гречишности», составляет в среднем 28%, что дает возможность рассчитывать на определенный эффект селекции по этому признаку.
Подобный анализ целесообразно проводить по каждому селекционному признаку. Так, мы подтвердили, что среднерусские пчелы при прочих равных условиях лучше посещают гречиху, чем пчелы других пород (Д.В.Гирник, Н.И.Кривцов, 1989). Дальнейшие исследования показали, что признак «гречишности» отличается достаточно высокой повторяемостью (табл. 3). Доля действия общих факторов, определяющих проявление «гречишности», составляет в среднем 28%, что дает возможность рассчитывать на определенный эффект селекции по этому признаку.
Корреляция. Основы учения о корреляции заложили еще Ч.Дарвин и Ж.Кювье. Ч.Дарвин (1859) утверждал, что если человек упорно ведет отбор по какому-либо определенному признаку и изменяет его, то в силу таинственных законов корреляции он почти наверняка вызовет изменения и других признаков. Ж.Кювье (по И.И.Шмальгаузену, 1969) писал: «Так как все органы животного образуют единую систему, части которой зависят друг от друга и действуют и противодействуют одна по отношению к другой, никакое изменение не может обнаружиться в одной части без того, чтобы не вызвать соответствующего изменения во всех остальных частях». Следует отметить, что корреляция изучена в пчеловодстве наиболее хорошо. Практически все научные статьи и диссертации изобилуют материалом с привлечением этого показателя. Хотелось бы обратить внимание исследователей (особенно молодых) на то, что не следует останавливаться на констатации факта найденной корреляции. Селекционный смысл этого параметра заключается в поисках сигнальных признаков для косвенного отбора. Ведь именно наличие корреляций открывает возможности для него. Расчет и изучение корреляций позволяет проводить раннюю оценку, давать прогноз развития признаков, по которым идет селекция.
Покажем целесообразность использования корреляций при оценке качества маток. Она укладывается в следующую схему косвенного отбора: выбраковка неполноценных маточников, отбор неплодных маток по оптимальной массе и размерам 3-го тергита и стернита, контроль за количеством оставшегося корма в маточнике после выхода матки (его отсутствие может указывать на недостаточность кормления), контроль за внешним видом маток, а в конечном счете их яйценоскости. Эта цепочка состоит из звеньев косвенного отбора, базирующегося на корреляции признаков. Подобные схемы можно выстроить и по другим признакам, прямая оценка которых затруднена.
К сожалению, практически все имеющиеся в настоящее время данные касаются фенотипической корреляции, представляющей собой параллельную изменчивость признаков под влиянием комплексного воздействия генетических факторов и факторов внешней среды. Требует изучения в пчеловодстве и генотипическая корреляция — параллельная изменчивость признаков под влиянием плейотропного действия генов или действия совместно наследующихся сцепленных генов, а также физиологическая корреляция, когда изменение одного признака обусловливает наличие другого.
Таким образом, весь селекционный процесс в пчеловодстве следует сопровождать оценкой генетико-популяционных параметров селекционных признаков, в большинстве своем являющихся полигенными. Без такого анализа селекция пчел будет неэффективной и бесплановой.
Н.И.КРИВЦОВ
НИИ пчеловодства, Рязанская обл., г. Рыбное

Феномен дупла
сен 27, 2022
Прополис и клещ Varroa destructor…
июнь 27, 2015
Лечебно-профилактические работы на пасек…
апр 3, 2019
Аморфа кустарниковая
фев 21, 2016
Экстенсивное пчеловодство…
апр 15, 2014
Медоносные растения севера Приамурья…
нояб 19, 2022
К чему может привести борьба с роением (…
окт 27, 2016
Работают две матки
мая 1, 2021
Заметки о прошедшем сезоне…
янв 1, 2016
Почему гибнут пчелы
дек 22, 2023
Рамки, рамки, рамки... (3)…
янв 17, 2018
Кулинария для пчел
сен 22, 2014
Хороший зимовник — залог успеха…
фев 3, 2017
Можно ли хранить мед в холодильнике?…
мая 14, 2014
Улей для Сибири
окт 24, 2023







 Адрес редакции журнала "Пчеловодство":
Адрес редакции журнала "Пчеловодство":