На страницах журнала «Пчеловодство» неоднократно обсуждалась роль естественной (тепловой) конвекции в ульях (особенно в зимних условиях). При этом обычно авторы давали различного рода оценки, но решения нелинейных уравнений конвекции не приводили.
На страницах журнала «Пчеловодство» неоднократно обсуждалась роль естественной (тепловой) конвекции в ульях (особенно в зимних условиях). При этом обычно авторы давали различного рода оценки, но решения нелинейных уравнений конвекции не приводили.
В данной работе кратко обсуждены результаты решения нелинейных уравнений свободной конвекции в ульях, впервые представленные на Международном съезде по тепловой конвекции (S.S.Kruglikov, E.L.Tarunin, 2003). Для расчета конвективного движения использовали уравнения свободной конвекции в приближении Буссинеска (Г.З.Гершуни, Е.М.Жуховицкий, 1972): сжимаемость воздуха учитывается лишь в члене с подъемной архимедовой силой. Рассчитывали двумерное течение в центральной части улья, ограниченной рамками с сотами.
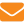
 Соответствующая область хорошо аппроксимируется так называемой ячейкой Хеле-Шоу (Г.З.Гершуни, Е.М.Жуховицкий, А.А.Непомнящий, 1988; Е.Л.Тарунин, 2003). Вид ячейки представлен на рис. 1; справа — сечение (область, занятая пчелами, заштрихована).
Соответствующая область хорошо аппроксимируется так называемой ячейкой Хеле-Шоу (Г.З.Гершуни, Е.М.Жуховицкий, А.А.Непомнящий, 1988; Е.Л.Тарунин, 2003). Вид ячейки представлен на рис. 1; справа — сечение (область, занятая пчелами, заштрихована).
Расчеты проводили при следующих параметрах ячейки: L=43,5 см, Н=30 см, D ≤ 2 см. Предполагалось, что пчелы, выделяющие тепло мощностью Q ≤ 10 Bт, расположены внутри шара с радиусом R. Плотность внутренних источников тепла вычисляли делением Q на объем шара радиусом R. Положение центра клуба задавалось параметрами a, b (рис. 1). Полагали, что центр клуба в основном расположен в центре улья.
Параметры воздуха (плотность, вязкость, теплоемкость, теплопроводность) брали при t = 20°С. Температуру внешнего воздуха считали равной нулю (t0=0). На узких гранях ячейки учитывали теплообмен по закону Ньютона: æ(dt/dn) = –α(t – t0) с параметром α ≤ 50 (Дж/m·c·ºK). Теплообмен с соседними ячейками не учитывали.
Вычисления выполняли в переменных функции тока, вихря скорости и температуры с порядком аппроксимации стационарного решения Q(hx2+hy2) на пространственной сетке 24x24 (Е.Л.Тарунин, 1990). Стационарное состояние с хорошей точностью достигалось за 7,5 мин (начальное состояние соответствовало покоящемуcя воздуху с нулевой температурой).
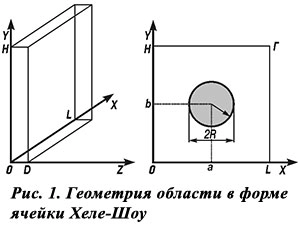
 Пример картины стационарного течения (изотермы слева и линии тока справа) представлен на рис. 2 для ячейки, проходящей через центр клуба. Изолинии проведены через равные интервалы. Ширина свободного зазора межрамочного пространства D = 1 см.
Пример картины стационарного течения (изотермы слева и линии тока справа) представлен на рис. 2 для ячейки, проходящей через центр клуба. Изолинии проведены через равные интервалы. Ширина свободного зазора межрамочного пространства D = 1 см.
Полная мощность тепловыделения клуба пчел в форме шара (R=14 см) Q = 4 Bт (в статьях, опубликованных ранее в журнале, указаны значения Q от 2 до 10 Вт), а параметр теплообмена α = 15.
По картине линий тока видно, что воздух поднимается по центру ячейки и опускается по краям с явными признаками пограничного слоя. Максимум скорости в центре улья 3,4 см/с (максимальное значение скорости около 4,4 см/с достигается в пограничном слое). Cредняя скорость ≈ 0,8 см/с.
Средняя температура воздуха в ячейке ≈15,7°С; максимальное значение 22,7°С отмечается в точке с координатами x = L/2, у ≈ 0,9 Н. По картине изотерм отчетливо видно, что в середине улья и нижней его части образуется устойчивая стратификация плотности воздуха, соответствующая расположению теплового воздуха сверху. Этот факт, возможно, не учитывали исследователи, которые полагали, что максимум разности температуры в улье можно измерить по разности температуры в центре и на границах.
Согласно расчетам разница между показателями температуры в центре улья и в его верхней части около 1,3°С, а разность между максимальными значениями в 3 раза больше. То есть тепловой поток на верхней границе улья больше такового потока на нижней границе в 3,4 раза.
Пользуясь рекомендациями Е.Л.Тарунина (2003), в модели ячейки Хеле-Шоу мы учитывали вязкое сопротивление на узких гранях (во многих работах этим эффектом пренебрегают). Если его не принять во внимание, на узких гранях максимальное значение скорости увеличивается примерно на 5%.
В то же время мы не учитывали возрастания вязкого сопротивления в области, занятой пчелами. Этот эффект в использованной модели частично погашается благодаря тому, что при расчетах мы берем ячейку с меньшей толщиной. Напомним, что представленные расчеты, рассуждения и выводы сделаны для центрального разреза улья, в других разрезах естественная конвекция значительно слабее.
В дальнейшем мы планируем выполнить расчеты, полагая, что область, занятая пчелами, — пористая среда или область с уменьшенным значением толщины ячейки. Кроме того, планируем учесть теплоизоляционный слой стенок улья и влияние влажности.
Е.Л.ТАРУНИН, С.С.КРУГЛИКОВ

Павильонные летковые заградители…
сен 26, 2014
Ловушки для роев
окт 3, 2020
Ольха — ранний пыльценос…
март 26, 2017
Распрямление проволоки…
янв 19, 2019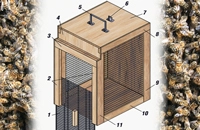
Универсальная роевня
июнь 5, 2023
Приспособление для отлова маток…
июнь 3, 2016
Куда исчезает вода? (окончание)…
март 22, 2015
Влияние стимулирующих подкормок на прием…
апр 29, 2023
Графическая интерпретация закономерности…
апр 13, 2015
Весенняя подкормка пчел…
март 15, 2015
Зима. Декабрь
дек 1, 2018
Естественная конвекция в ульях…
сен 29, 2018
Управление жизнедеятельностью пчел с пом…
июль 16, 2023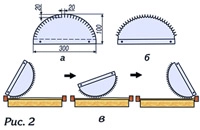
Натягивание проволоки и наващивание рамо…
июнь 10, 2016
В поисках оптимальных конструкций…
нояб 2, 2014









 Адрес редакции журнала "Пчеловодство":
Адрес редакции журнала "Пчеловодство":