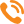Современный уровень знаний по физиологии терморегуляции в скоплениях медоносных пчел (зимний клуб) недостаточен для объяснения основных принципов жизнедеятельности пчелиной семьи в целом при низких температурах. До сих пор нет единого мнения по поводу регулируемой переменной, структуры и функции оболочки клуба, роли отдельных пчел при агрегировании и т.д. Исследователи едины лишь в том, что изоляция теплового центра внешним слоем пчел — решающий фактор в выживании при низких температурах.
Современный уровень знаний по физиологии терморегуляции в скоплениях медоносных пчел (зимний клуб) недостаточен для объяснения основных принципов жизнедеятельности пчелиной семьи в целом при низких температурах. До сих пор нет единого мнения по поводу регулируемой переменной, структуры и функции оболочки клуба, роли отдельных пчел при агрегировании и т.д. Исследователи едины лишь в том, что изоляция теплового центра внешним слоем пчел — решающий фактор в выживании при низких температурах.
Большинство моделей терморегуляции — это математические модели зимнего клуба пчел (S.Omholt, 1987; J.Watmough, S.Camazine, 1995; D.Sumpter, D.Broomhed, 2000; В.А.Тобоев, 2006), причем в них в основном моделируется не вся система терморегуляции, а лишь ее пассивная часть, охватывающая только стационарные режимы. Несмотря на очевидные их преимущества, нельзя недооценивать и теплофизические модели. Обладая реальной массой, теплоемкостью и температурой, находясь в реальных условиях теплообмена, они более наглядны и понятны большинству биологов, не владеющих специальными математическими знаниями.
В данной работе предлагается теплофизическая модель зимнего клуба пчел с системой регулирования центральной и периферической температуры. Это обобщенная натурная модель процессов теплопродукции и теплоотдачи, происходящих в скоплении пчел при изменении внешней температуры. Исследование проведено с помощью тепловизора ИРТИС-2000 (см. В.А.Тобоев, ж-л «Пчеловодство» №10, 2006). Типичная картина распределения температур в скоплении пчел показана на рисунке 1.
На основе исследования динамических особенностей тепловой картины на поверхности скопления пчел в предлагаемую модель заложена гипотеза регуляции температуры в центре клуба и на периферии по ее выходу из определенной зоны. Поддержание температурного гомеостаза осуществляется двумя сходными регуляторами, настроенными на разные интервалы температур, значения которых можно задавать в зависимости от реализуемой модели. Комфортными для пчел взяты значения температур в центральной части гнезда 24...32°С (при отсутствии расплода). С его появлением этот диапазон изменится до 33,5...35,5°С (Е.К.Еськов, 1992).
Электрическая схема предлагаемой модели показана на рисунке 2. Система регулирования температуры теплового центра Р1 настроена на интервал 24...32°С (пассивный период). При выходе температуры за его пределы либо включается дополнительный нагреватель Rд, имитируя увеличение теплопродукции в пчелином клубе, либо резистор R0, что соответствует увеличению теплоотдачи клуба при перегреве. Аналогично работает регулятор периферической температуры, поддерживая ее значение в пределах 8...13,5°С. Мощность нагревателя Rп подбирали так, чтобы средний уровень метаболизма покоя колебался от 3 до 5 Вт.

В ответ на охлаждение включается нагреватель Rд, увеличивая общий уровень метаболизма до 10 Вт. Имитация реакции увеличения теплоотдачи выполняется включением резистора R0, который уменьшает мощность нагрева примерно на 25%. Значения мощностей подбирают, изменяя напряжение питания нагревателей и сопротивление резистора R0. (Питание нагревателей идет от стабилизированного источника питания.)
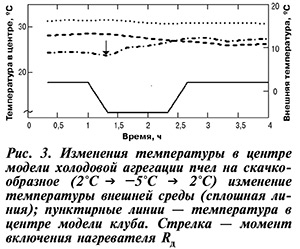 На рисунке 3 приведены характерные изменения температуры в центре модели зимнего клуба пчел в ответ на охлаждение окружающей среды. В зависимости от исходного теплового состояния модели возможно большое разнообразие в колебаниях температуры в центральной зоне, особенно в начальный период возникновения ответных реакций терморегуляции. Температура в центре может изменяться в направлении температурного стимула, оставаться на прежнем уровне или изменяться в противоположном направлении. При возврате окружающей температуры к исходному уровню температура в центре модели может вернуться к первоначальному уровню или остаться на новом установившемся. Это зависит от величины скачка внешней температуры, времени холодового воздействия и первоначального теплового состояния модели клуба. При этих воздействиях флуктуация температуры в различных точках модели приводит к изменению средней температуры, энергетической мерой которой может служить изменение теплосодержания.
На рисунке 3 приведены характерные изменения температуры в центре модели зимнего клуба пчел в ответ на охлаждение окружающей среды. В зависимости от исходного теплового состояния модели возможно большое разнообразие в колебаниях температуры в центральной зоне, особенно в начальный период возникновения ответных реакций терморегуляции. Температура в центре может изменяться в направлении температурного стимула, оставаться на прежнем уровне или изменяться в противоположном направлении. При возврате окружающей температуры к исходному уровню температура в центре модели может вернуться к первоначальному уровню или остаться на новом установившемся. Это зависит от величины скачка внешней температуры, времени холодового воздействия и первоначального теплового состояния модели клуба. При этих воздействиях флуктуация температуры в различных точках модели приводит к изменению средней температуры, энергетической мерой которой может служить изменение теплосодержания.
Реакция модели агрегации пчел на холодовое воздействие зависит от начального уровня теплосодержания. Как его повышение, так и снижение на определенную пороговую величину приводят к включению терморегуляторов, приводящих к отдаче тепла или его сохранению. Поэтому степень напряжения уровня терморегуляции определяется теплопродукцией qt, с учетом теплоотдачи qs и изменением теплосодержания Δq зимнего скопления пчел
qt = qs + Δq. (1)
Изменение теплосодержания в данной ситуации равно изменению количества тепла в пчелином клубе
Δq = mccpΔTcp, (2)
где m — масса пчел в клубе, кг; ccp — средняя удельная теплоемкость клуба, ккал/(кг°С) ; ΔTcp — изменение средней температуры клуба, °С.
Анализ специфики работы модели позволяет объяснить ряд противоречивых тенденций в поведении пчел при терморегуляции гнезда. Функционирование пчелиного клуба согласно данной модели направлено на сохранение такого соотношения между qt и qs, когда Δq ≈ const при определенных температурах. Другими словами, в определенном интервале температур (зона оптимальности — В.А.Тобоев, 2006) должно складываться динамическое равновесие между теплопродукцией и теплоотдачей, обеспечивающее соответствующие и относительно постоянные для каждого периода зимовки температуры внутри клуба и на его поверхности. Теплопродукция и теплоотдача выступают в качестве регулирующего воздействия по отношению к теплосодержанию. Вне зоны оптимальности теплосодержание клуба, определяемое прежде всего геометрией клуба и массой пчел в нем, претерпевает весьма заметные колебания вследствие неравенства вышеуказанных параметров. Равенство qt = qs справедливо лишь для интегральных величин теплообразования и теплоотдачи, взятых за достаточно большой промежуток времени. Что касается состояния клуба в какой-то момент, то равенство этих величин — редкое исключение. Следует отметить, что изменение теплосодержания может происходить под действием не только внешних, но и внутренних причин, например: при нарушениях режима влажности, увеличении концентрации углекислого газа в тепловом центре клуба, при патологических изменениях.
Таким образом, рассмотренная теплофизическая модель позволяет сделать вывод о том, что теплосодержание — интегральная величина, отражающая тепловое состояние пчелиного клуба. В этом случае система терморегуляции управляет уровнем теплонакопления в скоплении пчел, а не регулирует непосредственно температуру в определенной точке.
В дальнейшем на теплофизической модели зимнего скопления пчел предполагается провести ряд экспериментов с целью разрешения многочисленных вопросов, связанных со структурой и функцией системы терморегуляции пчелиной семьи как целостного организма.
В.А.ТОБОЕВ
Чувашский государственный университет

История разделительной решетки…
март 19, 2018
Анализ гетерогенности искусственных попу…
нояб 12, 2016
Опыты с летками
июнь 26, 2014
Улей с унифицированным основным корпусо…
дек 29, 2014
Горелка
мая 30, 2015
Про аскосфероз забыл
дек 15, 2014



Зольный щелок на моей пасеке…
окт 31, 2018
Павильон каркасной конструкции…
март 30, 2023
Пчелы и крылья
март 29, 2017
Необычное роение
окт 18, 2014
Весеннее наращивание пчел…
мая 27, 2022
Моя стационарная пасека…
апр 6, 2015








 Адрес редакции журнала "Пчеловодство":
Адрес редакции журнала "Пчеловодство":