В настоящем сообщении предпринята попытка оценки влияния температурного стресса в период раннего онтогенеза пчел на морфогенетические процессы, лежащие в основе снижения неспецифической устойчивости.
Медоносная пчела обладает широким диапазоном и уровнем естественной устойчивости, сформировавшимся в процессе эволюции вида. Это обстоятельство наряду с другими причинами позволило ей распространиться по всем континентам, за исключением Антарктиды. Однако до настоящего времени проблема устойчивости пчел к биотическим и абиотическим факторам внешней среды остается одной из ведущих проблем отечественных пчеловодов.
В наш век, несмотря на совершенство методов пчеловодства и интенсивное использование ветпрепаратов для профилактики и лечения вирусных и паразитарных болезней, гибель семей до и после зимовки сохраняется на высоком уровне. Естественно, возникает вопрос: почему же искусственно созданные популяции с генетически детерминированными и эволюционно закрепленными механизмами устойчивости в условиях интенсивного пчеловодства не могут реализовать свой наследственный потенциал?
От генотипа к фенотипу. При ответе на поставленный вопрос необходимо иметь в виду, что в процессе наследования происходит передача потомству не конкретных признаков или свойств, а лишь возможность их реализации. Эта возможность в ряде случаев определяется факторами внешней среды, то есть фенотип — результат взаимодействия генотипа и условий развития, среди которых температурный режим играет значительную роль. Классический пример влия-ния температурного режима в период раннего онтогенеза на степень выраженности признака приведен в работе Т.Моргана (Морган, 1919, цит. по Шмальгаузену, 1968). Длина крыльев у плодовой мушки дрозофилы с мутацией vestigial (короткие крылья) зависит от температурных условий выращивания личинок. При высоких температурах (32°С) у мух развиваются крылья почти нормальной длины. При низких температурах (18–25°С) крылья становятся практически рудиментарными.
Количественные изменения морфометрических признаков у пчел наблюдала М.Д.Еськова (2010). По ее данным, у пчел, развивавшихся при относительно высокой температуре, сокращались размеры тела и крыльев, а так же увеличивалась длина хоботка. Таким образом, температурный фактор способен влиять на «норму реакции», то есть степень выраженности признака, контролируемую генами.
Устойчивость — это генетически детерминированное свойство организма обеспечивать жизнеспособность при значительных неблагоприятных воздействиях. Она является результатом взаимодействия множества функционально связанных генов, объединенных в сложные иерархические системы генетических сетей, и факторов внешней среды.
Для формирования устойчивого фенотипа пчел исключительно важным является эмбриональный период развития. В это время отклонение температурного режима от оптимального приводит к нарушению целого ряда биохимических и морфогенетических процессов, и как следствие к снижению неспецифической устойчивости пчел.
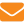
 Пчелиный расплод в условиях температурного стресса. Температурный стресс — это неспецифическая комплексная ответная реакция организма на экстремальные температурные условия развития, направленная на поддержание внутриклеточного гомеостаза. В отличие от взрослых особей, которые способны длительно выдерживать колебания температуры в широких диапазонах, пчелиный расплод проявляет особую чувствительность к температурным условиям существования. Оптимальная температура в расплодной части гнезда лежит в узком диапазоне — от 33 до 35°С. Оптимальной считается температура, при которой наблюдается наименьшая гибель появляющихся из расплода пчел. Оптимальный температурный диапазон оптимален как для развития рабочих пчел, так и для развития маток и трутней. По данным Е.К.Еськова (1995), при температуре 33–35°С до стадии имаго не доживает 2% расплода рабочих пчел, 4% расплода трутней и 6% расплода маток. Верхний предел температурной зависимости лежит на уровне 38–39°С, нижний – на уровне 28–29°С. За гранью указанных температур отмечается почти 100%-ная гибель расплода. Зависимость его гибели от температурных условий существования показана на графике. Таким образом, витальный температурный диапазон развития лежит в относительно узком диапазоне — 10°С.
Пчелиный расплод в условиях температурного стресса. Температурный стресс — это неспецифическая комплексная ответная реакция организма на экстремальные температурные условия развития, направленная на поддержание внутриклеточного гомеостаза. В отличие от взрослых особей, которые способны длительно выдерживать колебания температуры в широких диапазонах, пчелиный расплод проявляет особую чувствительность к температурным условиям существования. Оптимальная температура в расплодной части гнезда лежит в узком диапазоне — от 33 до 35°С. Оптимальной считается температура, при которой наблюдается наименьшая гибель появляющихся из расплода пчел. Оптимальный температурный диапазон оптимален как для развития рабочих пчел, так и для развития маток и трутней. По данным Е.К.Еськова (1995), при температуре 33–35°С до стадии имаго не доживает 2% расплода рабочих пчел, 4% расплода трутней и 6% расплода маток. Верхний предел температурной зависимости лежит на уровне 38–39°С, нижний – на уровне 28–29°С. За гранью указанных температур отмечается почти 100%-ная гибель расплода. Зависимость его гибели от температурных условий существования показана на графике. Таким образом, витальный температурный диапазон развития лежит в относительно узком диапазоне — 10°С.
Разные стадии развития расплода неодинаково реагируют как на колебания температуры в пределах витального диапазона, так и времени этого воздействия. Повышение температуры за пределы оптимума ведет к большей гибели расплода, чем понижение температуры.
Эмбриональная стадия. Из яиц, инкубируемых при оптимальной температуре и относительной влажности в пределах 71–82%, вылупляется около 95% личинок. При инкубации при 30°С гибель эмбрионов возрастает в 5–8 раз, при инкубации при 38°С — в 6–10. Однако краткая (1,5 ч) экспозиция яиц при 0°С ведет к гибели лишь 16% эмбрионов; остальные (где-то 80%) нормально заканчивают постэмбриональное развитие выходом пчел из ячеек.
Личиночная стадия развития. На личиночной стадии развития пчелиный расплод нуждается в частых контактах с пчелами, поддерживающими оптимальный температурный режим в улье. Как было установлено, при инкубации в искусственных условиях динамика гибели личинок в течение первых трех суток несущественно отличается при колебаниях температуры в диапазоне 7,5°С. От температуры существенно зависят изменения массы личинок: при 29°С за первые сутки масса тела уменьшалась в среднем на 2%, на 2-е — на 6,8% и на 3-и сутки — на 14% от нормы. При 34°С изменения были выше в среднем в 1,8 раза, при 37°С – в 2,5 раза. Гипертермия многократно сокращает продолжительность жизни личинок и предкуколок. Через сутки жизни при 29,5°С на стимуляцию током не реагировало 4% особей, на 2-е сутки — 6% и на 3-и сутки – 10% особей; при 37°С у 14, 20 и 39% особей соответственно отсутствовала реакция на стимуляцию током. Несмотря на то что время жизни некоторой части личинок значительно превосходило продолжительность личиночной стадии, ни одна из них не превратилась в куколку.
Колебания температуры на более поздних стадиях развития, например на стадии запечатанного расплода, хотя и не влияют на сам процесс органогенеза, тем не менее сопровождаются гибелью большого числа уже почти сформированных особей. Инкубация запечатанного расплода при 28°С приводит к элиминации 62% развивающихся пчел. Пчелы погибают, не предпринимая попыток выйти из ячеек, хотя и достигают того этапа развития, на котором возможна локомоторная активность.
Таким образом, все стадии доимагинального развития высоко чувствительны к колебаниям температуры за пределами оптимальной. При этом, как уже отмечалось, гипертермия ведет к более серьезным последствиям, чем гипотермия.
Температурный стресс как реакция адаптации. У всех животных переключение нормальной жизни клетки на стрессовую осуществляется на многих уровнях регуляции. Оно заключается в репрограммировании генома — тормозится экспрессия генов, активность которых характерна для жизни клетки в нормальных условиях, и активируются гены стрессовых белков (Жимулев, 2003). При воздействии высоких температур (мРНК — информационная РНК) кодирующие стрессовые белки обнаруживаются уже через 5 мин от начала стресса, а сами стрессовые белки отмечались в клетках уже через 15 мин после начала теплового шока. Их синтез активируется, достигая максимума за 2–4 ч теплового шока, а затем при снятии его начинает ослабевать, после окончания температурного воздействия синтез специфических белков прекращается, и возобновляется синтез белков, характерный для клетки при нормальных температурных условиях. При этом мРНК стрессовых белков быстро разрушаются в клетках, тогда как сами белки могут сохраняться существенно дольше, обеспечивая, по-видимому, повышение устойчивости клетки к нагреванию.
Длительное пребывание клеток в условиях теплового шока обычно также приводит к ослаблению и прекращению синтеза стрессовых белков. В этом случае включаются механизмы регуляции экспрессии генов стрессовых белков по принципу обратной связи.
Стрессовые белки представлены белками теплового шока (БТШ), низкомолекулярными белками теплового шока (нмБТШ), а также рядом ферментов (включая антиоксидантные ферменты). Главная функция БТШ состоит в правильной укладке вновь синтезированных полипептидов и в реукладке неверно уложенных и поврежденных полипептидных цепей (Жимулев, 2003).
В отличие от теплового шока холодовой шок в пределах положительных низких температур не вызывает денатурации белков, но ведет к изменению конформации молекул и изменению биохимических реакций. При холодовом стрессе происходит стабилизация вторичной структуры нуклеиновых кислот и, как следствие, ингибирование важнейших генетических процессов (ДНК-репликации, генной транскрипции и трансляции). Снижается активность многих ферментов и общего метаболизма, а также проницаемость мембран, что затрудняет транспорт веществ в клетку. Однако в клетках существует ряд механизмов, помогающих адаптироваться к гипотермии. Так, описан механизм, связанный с разобщением окисления и фосфорилирования в митохондриях. Воздействие низкотемпературного стресса также вызывает синтез специфических стрессовых белков, по аналогии с белками теплового шока, названных белками холодового шока (БХШ), которые вовлекаются в синтез белков и укладку мРНК.
Морфологическим проявлением генетической активности в ответ на повышение температуры на цитологическом уровне является формирование специфически активных районов хромосом — пуфов. Пуфы теплового шока на политенных хромосомах слюнных желез личинок дрозофилы образуются при повышении температуры с 20 до 37°С там, где они не появлялись при нормальной температуре. Такие же пуфы возникают в ответ на ферментные яды, блокирующие окислительное фосфорилирование. Реакция клеток на индуцирующие факторы очень быстрая. Формирование пуфов начинается уже через одну минуту после повышения температуры, в течение 20–30 мин они достигают максимальных размеров, а затем в течение нескольких часов регрессируют.
В последнее время обнаружена еще одна ответная реакция, возникающая под действием теплового и холодового шока, — активизация перемещений мобильных генетических элементов (МГЭ) (Пивоварова, Васильева, 2004). В экспериментах с некоторыми мутантными линиями плодовой мушки Drosophila melanogaster была показана активизация перемещений МГЭ под действием высоких и низких температур в период созревания мужских половых клеток (при спермиогенезе). Известно, что такое явление может сопровождаться генетическими эффектами, связанными с активацией и инактивацией генов, а также появлением структурных изменений хромосом. В силу этого генетические последствия теплового стресса могут проявляться в ряду следующих поколений.
Температурный стресс и жизнь пчелиной семьи. Приведенные выше данные неопровержимо свидетельствуют о том, что колебания температурного режима выше и ниже оптимальных значений вызывают серьезные изменения на молекулярном и цитологическом уровне, способных привести к гибели организма. Особенно чувствительными к колебанию температуры оказываются эмбриональные стадии, так как в это время происходят закладка всех органов и становление всех жизненно важных функций организма. Нарушение теплового режима нарушает временную согласованность работы эмбриональных генов, что и является одной из причин проявления летальности пчел, недоразвития органов и снижения показателей общей жизнеспособности отдельных особей. Все это, безусловно, сказывается на жизнеспособности пчелиной семьи в целом. Пчелы, пережившие тепловой или холодовой стресс, по всей видимости, обладают сниженными показателями жизнеспособности, сниженной способностью к полноценному выкармливанию потомства и сбору нектара. Систематические температурные стрессы, несомненно, снижают общую неспецифическую и специфическую устойчивость пчелиной семьи.
Приемы рационального пчеловодства — ранняя выставка на облет, раннее применение противороевых мер: деление семьи пополам, создание отводков, расширение гнезд, а также ранняя постановка магазинных надставок, частый осмотр пчел — ведут к нарушению теплового режима улья. Он изменяется также при частом и небрежном осмотре семей, переносе рамок, при избыточном открытии верхних и нижних летков в ранний весенний период. Последствия температурного стресса наиболее опасны в тот период, когда идет наращивание силы семьи. В летнее время аналогичные последствия испытывает семья при гипертермии. Особенно чувствительными к нарушению оптимального теплового режима оказываются слабые и средние по силе семьи. Именно они в полной мере испытывают все негативные последствия температурного шока, которые существенно ослабляют их силу к моменту медосбора и создают предпосылки для развития заболеваний. Нарушения теплового режима могут быть относительно быстро компенсированы только сильными семьями.
Все спасение — в сильных семьях – так считали классики пчеловодства.
М.А.МОНАХОВА
Биологический факультет
Московского государственного университета им. М.В.Ломоносова
И.И.ГОРЯЧЕВА
Учреждение Академии наук Институт общей генетики
им. Н.И.Вавилова РАН
Проведен анализ влияния по гипо- и гипертермии в эмбриональный и постэмбриональный период развития на жизнеспособность пчелиного расплода. Показано, что температурный стресс вызывает значительные изменения в структурно-функциональной организации генетической системы и экспрессии отдельных генов. Предполагается, что нарушение оптимальных температурных условий развития пчелиного расплода является одной из причин жизнеспособности, а также специфической и неспецифической устойчивости пчелы медоносной.
ЛИТЕРАТУРА
1. Еськов Е.К. Экология медоносной пчелы. — Рязань, 1993.
2. Еськова М.Д. Перегрев улья и развитие расплода // Пчеловодство. — 2010.
3. Жимулев И.Б. Пуфы теплового шока и синдром клеточного стресса. // Молекулярный механизм генетических процессов. — Новосибирск, 2003.
4. Пивоварова О.В., Васильева Л.А. Стрессовая индукция транспозиции ретротранспозонов mdg1 на разных стадиях сперматогенеза у самцов D. Melanogaster. // Экол. генетика. — 2004. — Т 2. — №3.
5. Шмальгаузен И.И. Факторы эволюции. Теория стабилизирующего отбора. — М.: Наука, 1968.
- ВКонтакте
- РћРТвЂВВВВВВВВнокласснРСвЂВВВВВВВВРєРСвЂВВВВВВВВ

Новый прием увеличения массы маток и тру…
дек 18, 2015
Трутни выделяют тепло …
март 28, 2024
Уход за пчелами в многокорпусном улье…
июль 16, 2015
Жить человеку в мире с пчелой…
нояб 4, 2016
Подготовка семей к зиме…
нояб 10, 2014
Апистимулин-А
фев 26, 2015
Акарасан при варроатозе и акарапидозе…
апр 11, 2015
Зимовка маток в нуклеусе…
фев 2, 2022
Заслон гепатиту и СПИДу…
март 30, 2015
Первая помощь при ужалении …
апр 18, 2024
Изменение профилей брусков…
март 1, 2020
Конкуренция между опылителями в медовом …
апр 30, 2015
Морфофункциональные особенности жилкован…
апр 26, 2023
Цветные трутни на пасеках Кировской обла…
мая 18, 2014
Селекция устойчивых к заболеваниям пчел …
мая 3, 2015








 Адрес редакции журнала "Пчеловодство":
Адрес редакции журнала "Пчеловодство":